肝细胞癌(HCC)是最常见的原发性肝癌类型,死亡率高,预后差。并且转移性HCC对治疗药物的耐药性更强。原因在于,在转移进展过程中,癌细胞可利用脂质为远处转移提供能量。所以,脂质代谢重编程实际上是一个癌症标志,它是各种因素控制的复杂级联反应,而AMP活化蛋白激酶(AMPK)和乙酰辅酶a羧化酶(ACC)是其中的两个关键调控因子。
环状RNA(circRNAs)是一类内源性RNA转录本,可在癌症中发挥调节作用。环状RNA也被报道在HCC中发挥作用。但目前还没有发现环状RNA通过调控脂质代谢来调节HCC的转移。另外,许多在癌症中具有确定功能的环状RNA在人类和小鼠之间并不保守,这在某种程度上阻碍了在动物模型中环状RNA潜在机制的深入研究。所以,鉴定在HCC中起关键作用的保守环状RNA,可以对潜在生物标志物和治疗靶点的挖掘提供更有意义的参考。
2023年11月12日,中国科学技术大学单革教授团队在Advanced Science(IF=15.1)发表文章”A Mammalian Conserved Circular RNA CircLARP1B Regulates Hepatocellular Carcinoma Metastasis and Lipid Metabolism”。在本研究中,一种哺乳动物保守的circRNA——circLARP1B可以在肝细胞癌(HCC)中发挥关键作用。circLARP1B在预后和总生存期较差的患者表达水平高,并可通过促进HCC中脂肪酸的合成,增强细胞脂质积累和癌症转移特性。在诱导的HCC模型中,敲低circLARP1B会减少癌症转移和脂质积累。具体机制在于,circLARP1B与细胞质中的异质核糖核蛋白D(HNRNPD)结合,降低LKB1 mRNA稳定性和蛋白水平,促进HCC转移和脂质积累。靶向circLARP1B敲低可作为HCC治疗的潜在靶点。

CircLARP1B作为一种哺乳动物保守的CircRNA,在HCC转移中被鉴定出来
RNA-seq检测6个HCC标本用于研究保守环状RNA在HCC中的功能作用。经过一系列的分析(表达水平/保守型分析),最终鉴定出circLARP1B是表达最高,也是唯一转移上调的circRNA,并且序列保守。circLARP1B由La核糖核蛋白1B(LARP1B)基因的外显子2-4组成。随后作者对circLARP1B进行了成环/耐受/定位胞质验证。
NCBI和circAtlas数据库分析的circLARP1B和LARP1B mRNA在人体组织中表现出不同的表达模式;例如,在甲状腺组织中表达LARP1B mRNA,而不是circLARP1B。LARP1B表达量较高和较低的两组HCC患者之间的总生存率无显著性差异,说明LARP1B mRNA可能不是HCC的关键调控因子。转移性HCC标本中circLARP1B水平高于非转移性标本。并且患者的HCC标本结果显示,较高水平的circLARP1B与转移的标本和TNM晚期(III-IV)以及生存率低密切相关。综上所述,circLARP1B作为一种哺乳动物保守的circRNA可能在HCC转移中发挥促进作用。
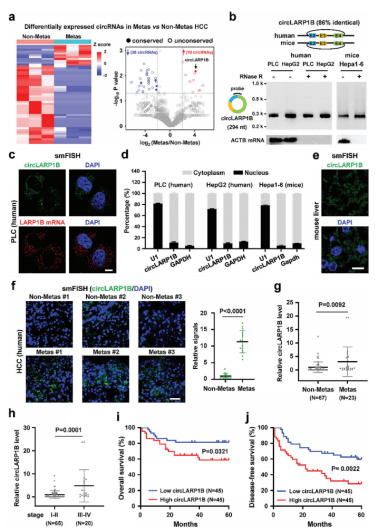
图1 哺乳动物保守的circLARP1B在HCC转移中的特征分析
circLARP1B介导脂肪酸合成调控细胞侵袭和脂质积累
环状外显子的侧翼内含子分别包含Alu元素和B1重复序列,属于反向互补序列,可以促进环状RNA的生物发生。为了探索circLARP1B的细胞功能,使用(CRISPR)/Cas9技术生成circLARP1B缺陷(circLARP1B-Def)PLC细胞。具体方法是,删除了LARP1B第4个内含子中包含4个近端Alu元件的重复序列。circLARP1B-Def PLC细胞的circLARP1B水平显著降低,而LARP1B mRNA水平没有变化。
Transwell侵袭实验发现,CircLARP1B-Def PLC细胞相对应的侵袭能力明显降低。不过在生长曲线和集落形成方面没有显著差异。侵袭能力与转移直接相关,生长曲线和集落形成与细胞的生长和增殖有关。然后,作者通过液相色谱-质谱(LC-MS)非靶向代谢组学评估了circLARP1B-Def PLC细胞中潜在的代谢改变。在6种受到显著影响的代谢途径中,脂质代谢受到的干扰最大。
然后,作者进行了脂质组学来检测脂类组成,检测到的24种失调脂类。在circLARP1B缺失下,这24种脂类发生了显著变化,表现在磷脂酰乙醇胺(PE)、双甘油酯(DG)、甘油三酯(TG)和磷脂酰肌醇(PI)降低。为了进一步探索circLARP1B促进脂质积累的机制,作者用13c标记的靶向代谢通量分析进行了代谢标记,以评估circLARP1B-Def和WT PLC细胞中13c标记的葡萄糖和谷氨酰胺掺入脂质的情况。靶向代谢通量分析结果显示,共检测到17种具有13C标记的脂肪酸,发现它们在circLARP1B-Def细胞中均显著降低,表明circLARP1B对脂肪酸合成(FAS)有促进作用。葡萄糖和谷氨酰胺首先转化为双碳乙酰辅酶a,乙酰辅酶a作为FAS中ACC1酶的底物。Unc-51样自噬激活激酶1(ULK1)是脂肪自噬起始的关键调控因子,ULK1的Ser555磷酸化诱导脂肪自噬。ULK1和磷酸化ULK1(p-ULK1)水平在circLARP1B-Def和WT细胞之间没有明显的变化。综上所述,这些发现表明,circLARP1B促进了FAS的脂质积累,而对脂肪自噬无明显作用。
ACC1的活性被Ser79的磷酸化所抑制,观察到在circLARP1B-Def细胞中,磷酸化的ACC1(pACC1)水平显著上调。AMPK是催化ACC1磷酸化的主要激酶。circLARP1B-Def细胞中磷酸化的AMPK(pAMPK)水平也显著高于WT细胞。而较高水平的pAMPK和p-ACC1会抑制circLARP1B-Def细胞的脂肪生成。AMPK的激活也会导致HCC中一系列代谢物的变化。通过查阅文献发现,12个circLARP1B调节的代谢物在HCC中受AMPK信号调控,其中实验发现,8个代谢物在circLARP1B缺失和AMPK激活之间表现出一致的变化。
此外,作者还研究了circLARP1B对其他信号通路(受AMPK调控)的影响,如mTOR通路、血管生成、TGF-β和线粒体活性。与WT细胞相比,circLARP1B-Def细胞AMPK激活导致mTORC1磷酸化,抑制mTOR通路,抑制脂肪酸合成。这提示,circLARP1B可调控AMPK直接调控的ACC1和mTOR信号通路,从而发挥功能。
因为ACC1受AMPK直接调控,是FAS的限速酶,接下来重点研究ACC1。敲低circLARP1B可抑制细胞侵袭和脂滴形成,并增加了细胞中的p-AMPK和p-ACC1的水平。而过表达则相反。脂肪酸合成酶(FASN)是一种人脂肪生成酶,在circLARP1BDef PLC细胞中表达上调。FASN过表达可增强circLARP1B-Def PLC细胞被抑制的细胞侵袭和脂滴形成能力。使用用强效FASN抑制剂IPI-9119处理过表达circLARP1B的PLC细胞,发现FASN抑制剂阻断了circLARP1B对脂滴形成和细胞侵袭的促进作用。综上所述,circLARP1B可在细胞水平上重塑脂质代谢来刺激HCC,并通过调节AMPK及其下游靶点ACC1来促进FAS。
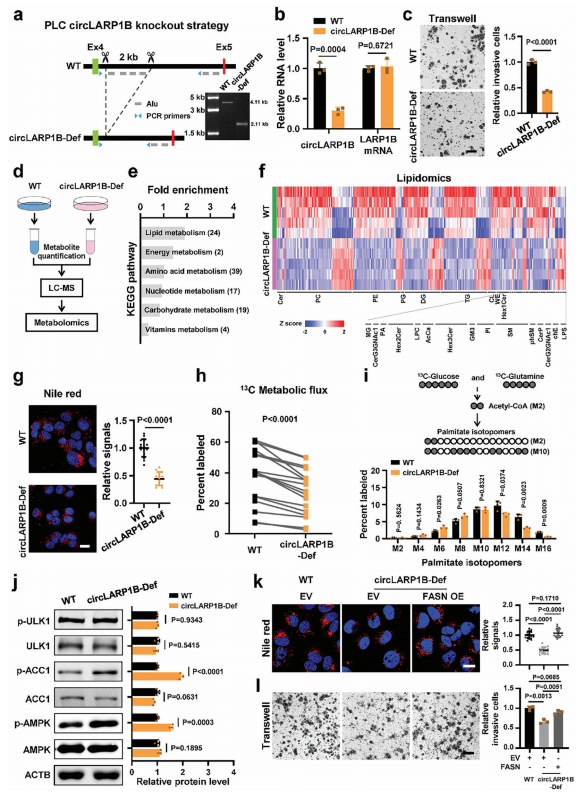
图2 circLARP1B介导脂肪酸合成调控细胞侵袭和脂质积累
circLARP1B与HNRNPD蛋白相互作用
作者解析circLARP1B潜在的相互作用分子来阐明其功能机制。AGO2的RNA免疫沉淀(RIP)解析靶circRNA和miRNA的相互作用,发现在PLC细胞中没有显示circLARP1B的富集,提示circLARP1B有限的miRNA结合能力。此外,核糖体谱分析,发现circLARP1B与多聚体没有明显的结合,提示circLARP1B有限的蛋白翻译能力。
不过作者使用生物素的探针对circLARP1B的BSJ进行RNA下拉,跑胶分离+银染质谱。HNRNPD,也被称为富含au的元素rna结合因子1(AUF1),被筛选为circLARP1的互作蛋白。随后作者又体外再度验证了这一结果。不过HNRNPD不结合LARP1B mRNA。
为了绘制circLARP1B中HNRNPD的结合位点,我们通过RBP map发现了人类和小鼠circLARP1B中保守的两个基序(基序1和基序2),这是一种预测RNA序列中RBP结合位点的工具。这两个位点也富au,与HNRNPD对富au区的结合偏好一致。circLARP1B中基序1或基序2的突变降低了circLARP1B和HNRNPD之间的相互作用,这而两个基序的双突变则直接消除了相互作用。
HNRNPD与mRNA结合可以调节RNA的稳定性,敲低HNRNPD并不影响circLARP1B的水平,表明HNRNPD并不调节circLARP1B的稳定性。反过来敲低circLARP1B也不影响HNRNPD mRNA和蛋白的水平。综上所述,具有两个功能位点的circLARP1B可以与大量的细胞质HNRNPD相互作用,但是circLARP1B和HNRNPD不相互调节对方的表达或稳定性。
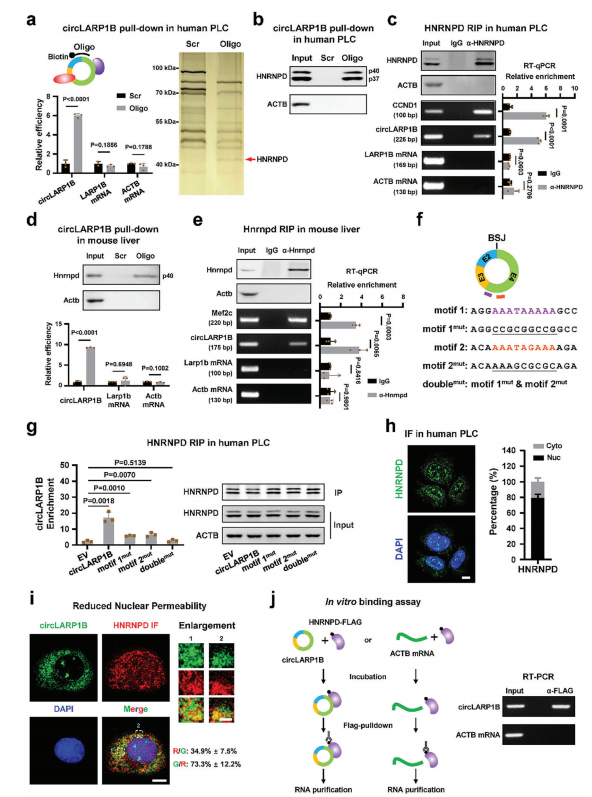
图3 circLARP1B与HNRNPD蛋白相互作用,但互不影响表达
circLARP1B结合和调节HNRNPD发挥功能作用
然后,作者开始研究HNRNPD结合在circLARP1B功能中的作用。circLARP1B过表达可以挽救circLARP1B-Def细胞被抑制的细胞侵袭和LD形成。而HNRNPD的过表达抑制了细胞侵袭和LD的形成。与HNRNPD一起过表达circLARP1B可阻断了HNRNPD对细胞侵袭、LD形成以及p-AMPK和pACC1水平的影响。综合这些结果提示,circLARP1B可能通过结合和调节HNRNPD发挥作用。
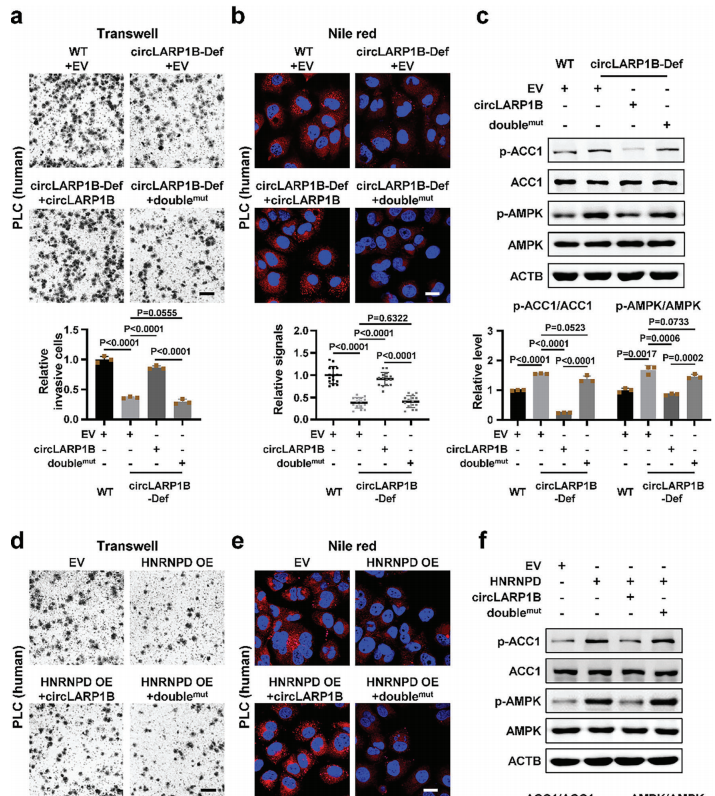 图4 circLARP1B结合和调节HNRNPD发挥功能作用
图4 circLARP1B结合和调节HNRNPD发挥功能作用
circLARP1B通过干扰HNRNPD降解LKB1 mRNA
为了进一步了解circLARP1B和HNRNPD在HCC细胞中的分子机制,作者进行HNRNPD RIP,对RIP的RNA进行RNA-seq。结果发现,mRNA靶标上的分布主要显示在3‘UTR,这与细胞质HNRNPD通过与3’UTR结合调节mRNA稳定性的功能作用一致。将来自circLARP1B-Def细胞和circLARP1B过表达细胞的RIP-seq数据与来自相应的对照细胞进行比较,发现受circLARP1B水平变化影响的mRNA子集可能在竞争HNRNPD时存在劣势,因为其与该蛋白的结合能力相对较弱(图5a)。
对这些靶基因的基因本体论(GO)发现,Thr磷酸化最显著(P值最低),其中LKB1也被称为丝氨酸/苏氨酸激酶11(STK11)在3‘UTR上显示出最全面的HNRNPD结合信号。众所周知,LKB1可以直接催化AMPK的Thr172的磷酸化,而这是AMPK激活所必需的。RIP-seq数据发现,circLARP1B缺失会促进HNRNPD结合LKB1,而circLARP1B过表达则相反。但circLARP1B过表达,HNRNPD结合circLARP1B多,反之则反。过表达的LKB1阻断了circLARP1B促进细胞侵袭和LDs形成的作用,也几乎消除了circLARP1B对p-AMPK和pACC1水平的影响。与非转移性HCC标本相比,转移性HCC标本中的LKB1蛋白水平较低。这些结果表明,LKB1 mRNA是circLARP1B的下游靶点,二者的功能作用相反。
circLARP1B减少,LKB1 mRNA的稳定水平增加。那HNRNPD和circLARP1B互作是否调控LKB1 mRNA的稳定水平呢?实验结果表明,敲低HNRNPD会导致细胞中LKB1 mRNA的半衰期和LKB1蛋白水平显著降低,表明HNRNPD正向调控LKB1 mRNA稳定。而在circLARP1B敲低却能增加LKB1 mRNA的稳定性和蛋白水平,过表达则相反。综上,circLARP1B虽然不影响HNRNPD表达,但会抑制LKB1表达,具体机制在于circLARP1B可以阻断HNRNPD对LKB1基因稳定的促进作用。
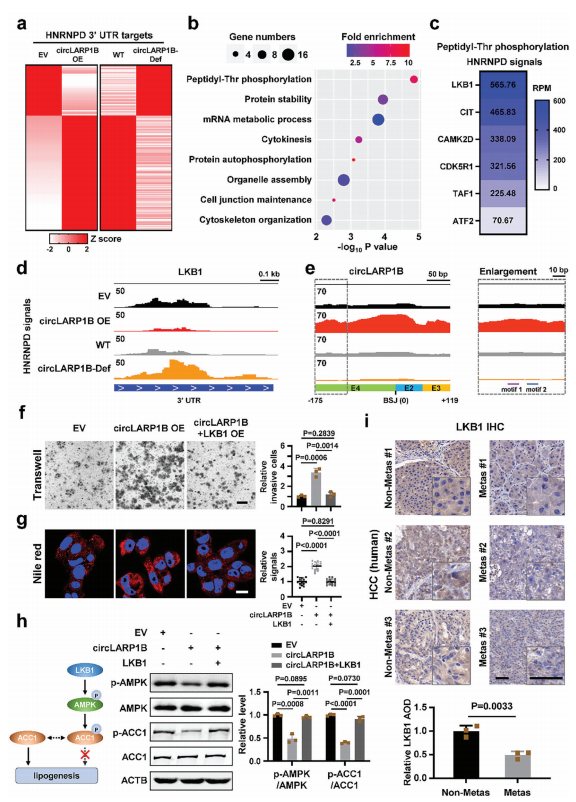
图5 circLARP1B通过干扰HNRNPD降解LKB1 mRNA
circLARP1B降低可抑制HCC进展
作者使用CRISPR/Cas9生成circLARP1B缺陷小鼠(circLARP1B−/−),诱导小鼠肝脏中circLARP1B的表达显著降低,而Larp1b mRNA和蛋白水平不变。通过油红O染色检测发现,circLARP1B敲低小鼠肝脏中的ld显著降低,而p-Ampk和p-Acc1的蛋白水平显著升高。
为了在机体水平上检测circLARP1B在HCC中的生理作用,作者使用二乙基亚硝胺(DEN)诱导小鼠HCC,DEN已被证实会导致严重的肝脏损伤并最终导致肝癌发生。在DEN诱导18周后(HCC小鼠),与WT小鼠相比,circLARP1B−/−小鼠的肝脏肿瘤淋巴结表现出更少、更小,肺转移淋巴结明显更小,抗转移标志物e-钙粘蛋白升高。在circLARP1B−/−HCC小鼠中,与HCC相关的生物标志物均有降低。综上所述,敲低circLARP1B水平能够抑制HCC的进展、脂质积累和转移。
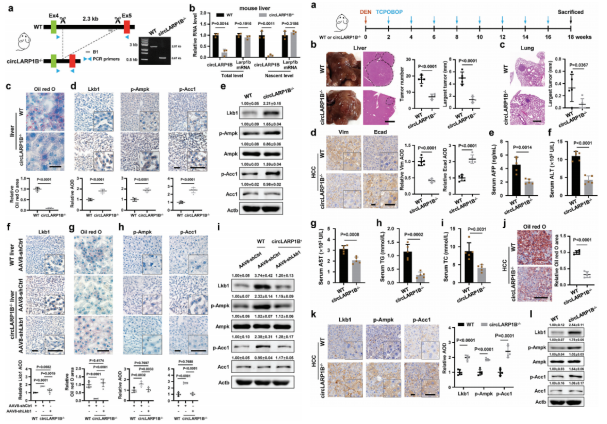
图6 circLARP1B降低可抑制HCC进展
LKB1是circLARP1B在HCC中发挥作用的关键靶点
敲低circLARP1B抑制HCC进程已有定论。circLARP1B−/−HCC小鼠模型显示,敲低Lkb1可以诱发HCC,表现为促进肝肿瘤增生和肺淋巴结转移以及Vim水平增高,e-钙粘蛋白升高降低,情况和WT的HCC小鼠一样。circLARP1B−/−HCC小鼠肝脏中观察到的p-Ampk和p-pAcc1水平的升高,Lkb1敲低后也被降低。这提示,Lkb1作为关键因子参与肝细胞中circLARP1B介导HCC脂质代谢和转移过程。
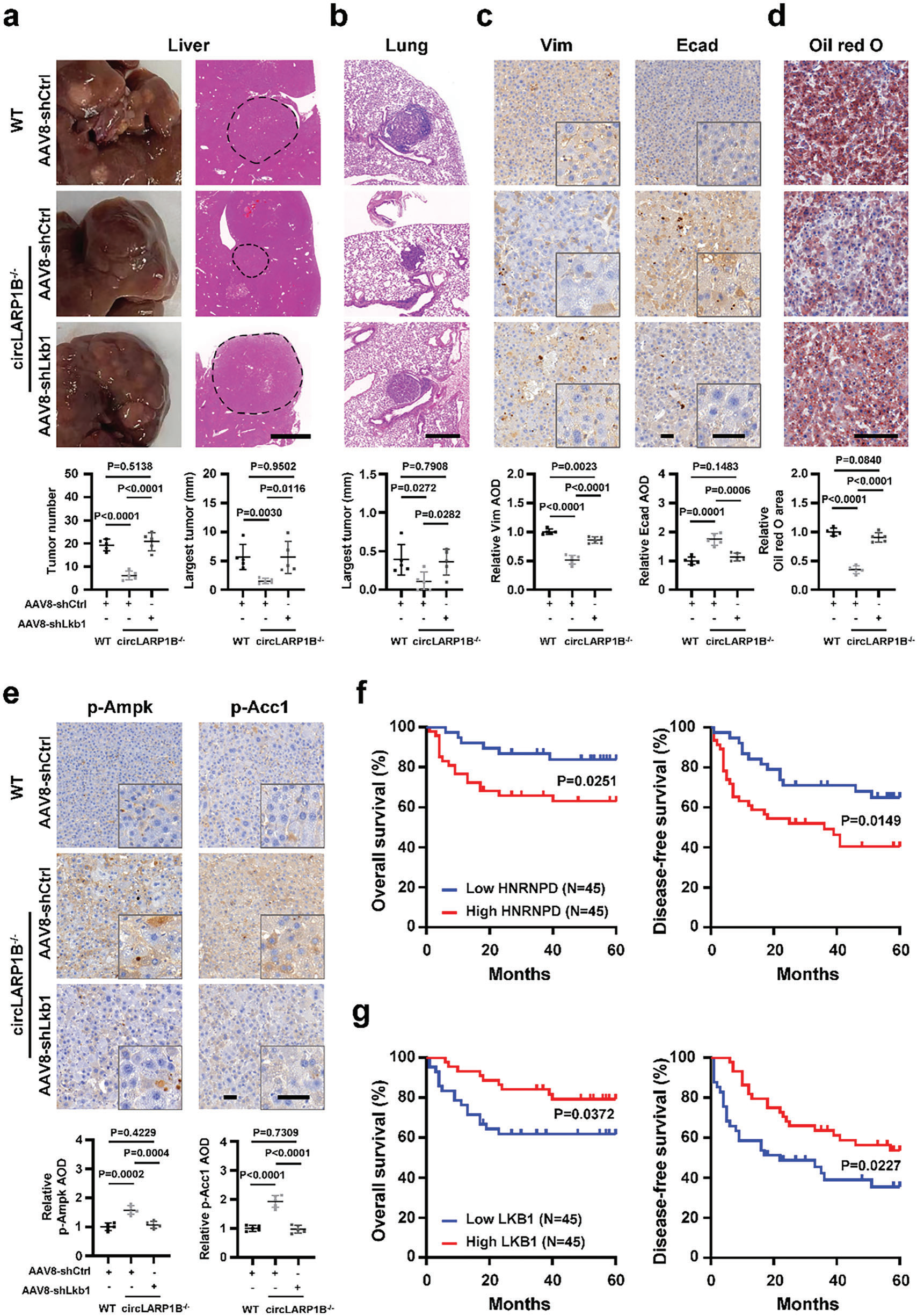 图7 LKB1是circLARP1B在HCC中发挥作用的关键靶点
图7 LKB1是circLARP1B在HCC中发挥作用的关键靶点
原文链接:
https://doi.org/10.1002/advs.202305902