RNA疗法已经发展成为一种强大的治疗策略,然而其较短的半衰期限制了效用。RNA环化可抵抗核酸外切酶介导的降解,被认为是一种减缓RNA降解的有前景的方法。环状RNA(cRNA)的生产通常涉及RNaseR富集和高效液相色谱(HPLC)纯化,以减少体外转录(IVT)过程中产生的双链RNA引发的免疫反应,因此产量较低(通常低于输入RNA的1%)。cRNA的高效生产和纯化方法,一定程度上限制了其更广泛的应用。
通过cRNA实现的用途涉及与免疫系统的相互作用。已有不少cRNA表达抗原诱导免疫应答用于疫苗开发的研究。然而,基于cRNA靶向基因组和表观基因组的疗法中,免疫反应可能会抑制治疗效果。因此,去免疫是一个更难解决的问题,尤其是在cRNA递送非人源蛋白的情况下。
近日,美国加州大学Prashant Mali研究团队在Nat Biomed Eng杂志上发表研究论文:Robust genome and cell engineering via in vitro and in situ circularized RNAs。论文阐述了研究团队开发的两种RNA环化方法:利用II组内含子体外制备cRNA (ocRNA),以及使用内源性普遍表达的RtcB蛋白在细胞内环化生成cRNA (icRNA)。研究还证实了cRNA可应用于干细胞工程以及通过锌指(ZF)蛋白和CRISPR-Cas9系统实现强大的靶向基因组和表观基因组作用。

一、 制备ocRNA和icRNA
研究采用来自破伤风梭菌的II型内含子,并添加侧翼扭转核酶改善远程相互作用,体外制备ocRNA。通过Sanger测序检测外显子结合位点(EBS)之间的26nt剪接痕,证实了RNA环化。进一步分析环化产物实现环化效率在70%左右,纯化后产率60%。(图1)
此外,研究通过加入在目的序列上IRES和有效负载的两侧使用扭转核酶,转录出线性RNA,递送进细胞后,在普遍存在的细胞内源性RNA连接酶RtcB作用下,线性RNA连接5 ‘和3 ‘端,实现原位环化。icRNA环化水平~20%,随时间推移比例增加,且cRNA表达的GFP比线性原位环状化缺陷RNA (icdRNAs)的持久。(图1)
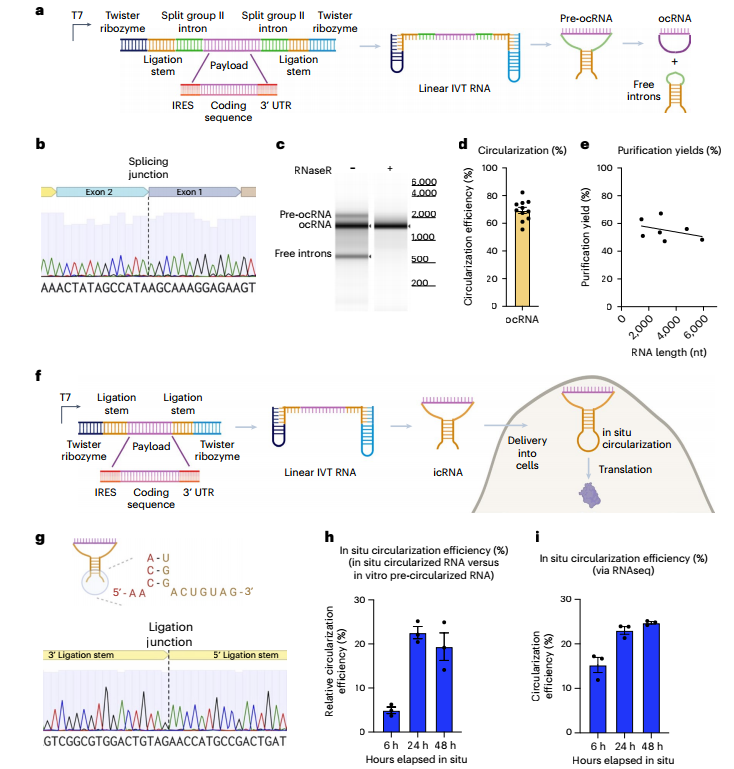
图1 制备ocRNA和icRNA
二、工程改进cRNA的翻译
研究筛选了19个天然存在和合成的IRES序列,发现脑心肌炎病毒IRES(EMCV)和柯萨奇病毒B3 IRES介导cRNA翻译效果最好,因此加入EMCV IRES元件及有助改善翻译的3’UTR和poly(A) stretch重新设计cRNA,以提高cRNA的翻译效率。(图2)
三、开发简易的cRNA纯化方案
为寻求使用更简单和更经济的纯化方法,在icRNA IVT过程中加入尿素作为变性剂,减少通过T7聚合酶转录所固有的高度免疫原性和dsRNA产生;对于ocRNA,因为在添加变性剂后,剪接的关键二级结构被取消,因此,依赖纤维素色谱法选择性去除长度超过30 bp的dsRNA(长于cRNAs中关键的IRES二级结构)。纯化后的icRNA和ocRNA表现出轻微的免疫反应,ocRNA诱导的免疫翻译几乎无法检测到。虽然HPLC纯化是临床翻译的首选方法,但上述cRNA纯化方法为大多数体外和体内应用提供了可行的替代方法。(图2)
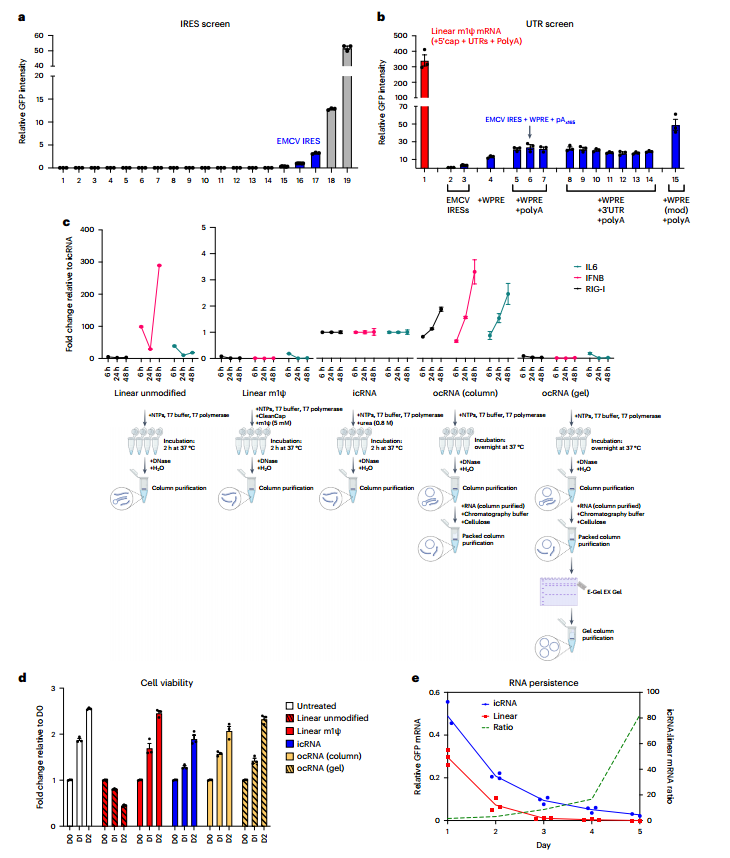
图2 ocRNA和icRNA的优化与表征。
四、cRNA的功能表征
利用干细胞来源的神经元细胞和心肌细胞验证cRNA的表达,结果发现与编码GFP的线性RNA相比,cRNA表达明显更持久。进一步结果表明,潜在细胞内容物的表达取决于IRES活性和/或对非分裂细胞的偏好,与分裂细胞相比,非分裂细胞在转染后不会迅速稀释RNA水平。H1人类多能干细胞(hPSCs)中单次转染NEUROD1 cRNA,可在1周内分化为神经元,这证明了干细胞向非分裂细胞的分化,可以更好地利用持久icRNA提供的高表达的蛋白质积累。(图2-3)
研究进一步制备icRNA或icdRNA的脂质纳米颗粒(LNPs),眼球后静脉注射到小鼠体内,结果表明icRNA可在体内原位环化,cRNA在小鼠体内至少可持续表达7天,而线性mRNA在第7天后大部分降解。
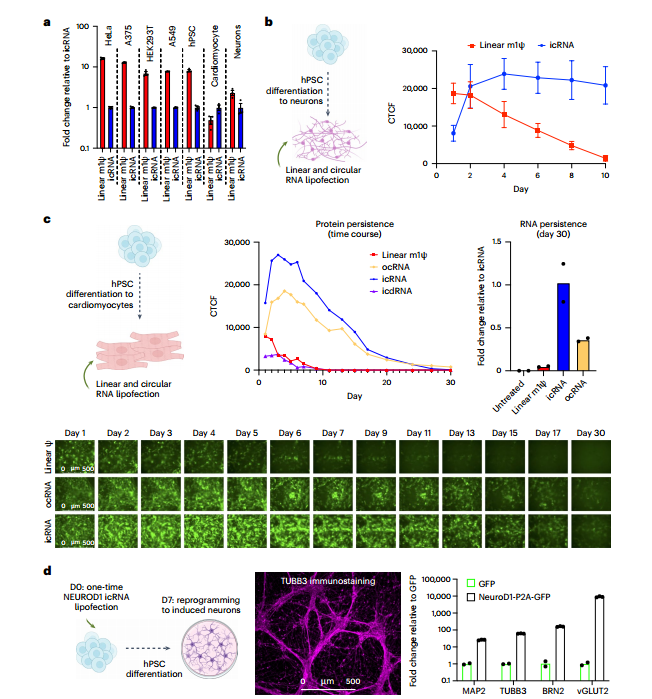
图3 评估ocRNA和icRNA的持久性和活性。
五、cRNA递送ZF蛋白实现基因组和表观基因组工程
除了持续转基因表达的应用之外,cRNA还可能促进有效靶向基因组,特别是表观基因组的应用。ZF蛋白作为一种完全基于蛋白质的基因组工程,特别适合cRNA这种持续的传递模式。研究证明,锌指核酸酶(ZFN) icRNAs确实可以进行更有效的基因组编辑。(图4)
PCSK9的功能缺失突变与心血管疾病风险降低相关。研究在HeLa细胞中筛选出16个靶向hPCSK9转录起始位点,有效抑制基因表达的ZF-KRAB蛋白模块;利用cRNA融合表达3A3L DNA甲基转移酶和ZF-KRAB蛋白模块,在12天内有效抑制了PCSK9的表达;利用cRNA递送ZF瞬时表达表观基因组调控子,可以实现强效的基因抑制,从而实现无基因组疤痕且安全的方法调节治疗性基因表达。(图4)
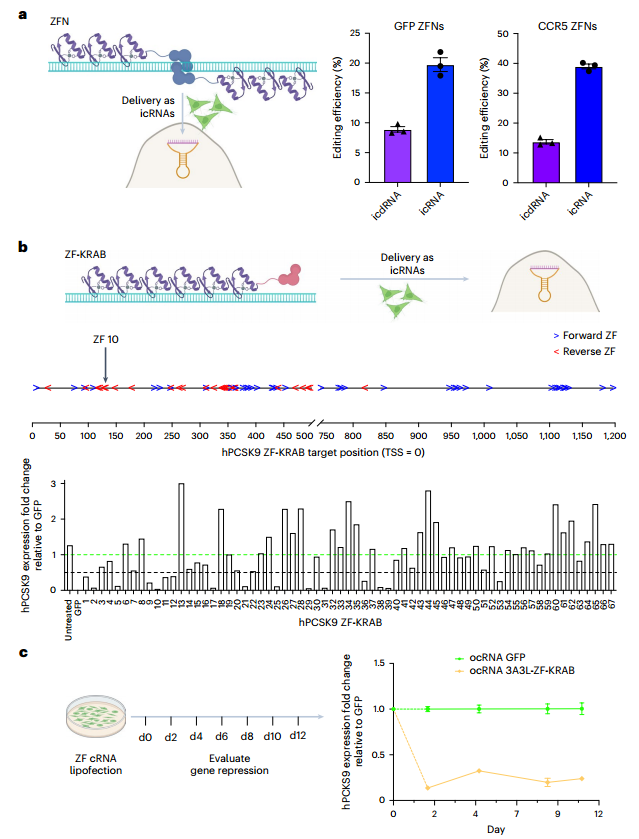
图4 icRNA和ocRNA在ZF介导的基因组和表观基因组靶向中的应用
六、通过cRNAs递送去免疫CRISPR-Cas实现基因组和表观基因组工程
因为CRISPR系统来自原核生物,cRNA的持久性表达特征可能会加剧CRISPR系统在治疗环境中的免疫反应。因此,在维持cRNA持久性表达的情况下,有必要通过组合突变,尤其是突变具有免疫原性的表位,筛选去免疫Cas9蛋白。研究开发了一种能筛选百万个基因的LORAX蛋白工程方法,在自然环境中检测核苷酸多态性(SNPs),筛选突变文库。研究最终结合netMHC表位预测软件对免疫原性的评估,确定导致免疫原性表位去免疫的最有效候选突变。(图5)
然后,利用长读Nanopore测序检测组合突变文库的筛选结果,Nanopore测序很好地规避了短读测序的限制,对整个目标基因进行单分子测序,使文库设计策略能够覆盖分子的全长区域。最后,研究经过一系列的筛选和验证得到功能正常的去免疫Cas9。(图5)
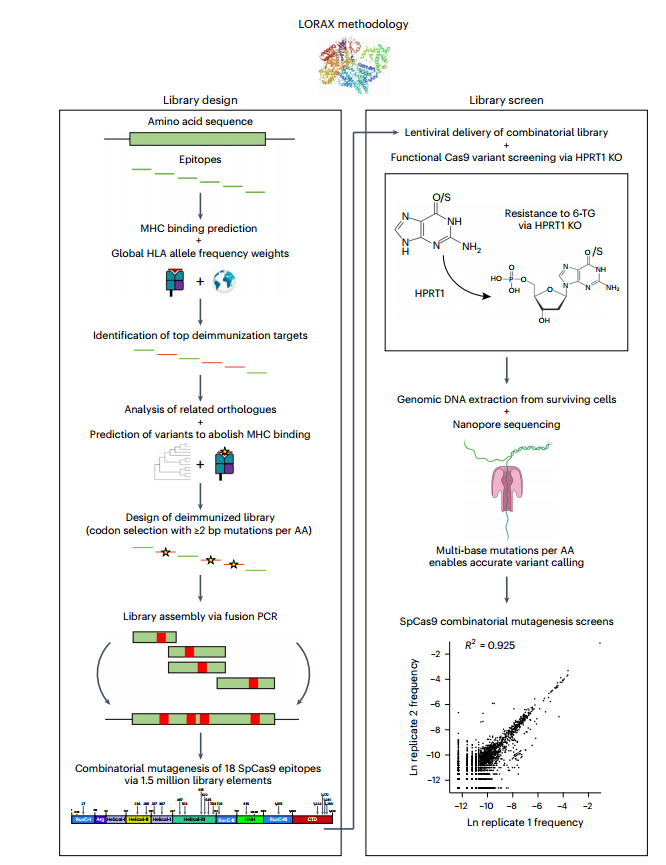
图5 LORAX蛋白工程方法筛选去免疫Cas9变异体。
研究将WT SpCas9或突变体SpCas9v4的icRNA,以及靶向AAVS1的sgRNA转染到HEK293Ts中,成功编辑AAVS1基因组;将WT型dSpCas9或突变型dSpCas9v4 CRISPRoff的icRNA和ocRNA以及靶向B2M基因的sgRNA转染到HEK293Ts中,证实了对B2M基因的抑制作用。通过测试分析发现,体外环化的ocRNA对SpCas9和CRISPRoff插入序列的环化效率分别约为40%和20%。研究通过RNA-seq证实了SpCas9v4和SpCas9在该实验中具有相当的特异性。(图6)
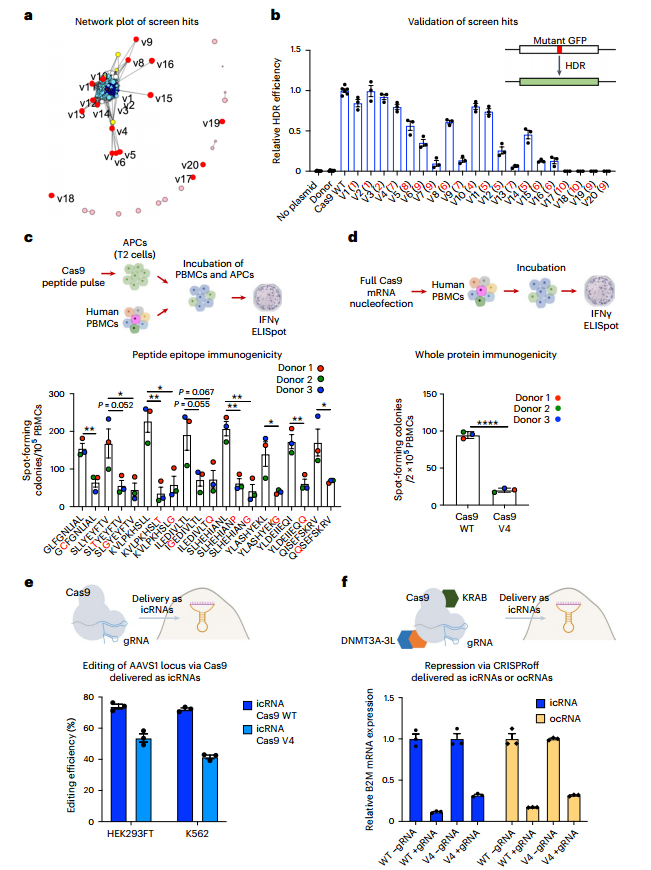
图6 LORAX筛选验证鉴定出Cas9去免疫变异体,以及通过icRNA和ocRNA递送靶向基因组和表观基因组作用。
总结
研究开发了利用II型内含子体外环化cRNAs,还通过内源表达的RtcB蛋白进行细胞内原位环化产生cRNA。此外,研究开发了无高效液相色谱纯化方案,使cRNA高产量,且保持低免疫反应。由此产生的cRNA简单和可扩展地促进了干细胞工程,以及通过ZF蛋白和CRISPR-Cas9系统进行强大的靶向基因组和表观基因组的一系列应用。特别是,携带EMCV IRES的icRNA和ocRNA在心肌细胞和神经元中表现出强大的表达(与线性加帽和修饰的RNA相比),并且延长了RNA的持久性,强调了cRNA在非分裂细胞中的巨大前景和效用。此外,研究团队的环化策略允许高效地生成和递送大分子蛋白,如Cas9和CRISPRoff,有效解决慢病毒和腺相关病毒包装限制的问题。
原文链接
https://www.nature.com/articles/s41551-024-01245-z