胖志
2020年3月14日,中山大学肿瘤医院符立梧和新乡医学院王现伟为共同通讯作者在Molecular Cancer (IF=10.679)杂志上发表了一篇题为“Functions and mechanisms of circular RNAs in cancer radiotherapy and chemotherapy resistance”的综述,总结了circRNA在肿瘤放化疗耐药中的最新研究进展及其机制,并讨论了现有知识的局限性和潜在的未来发展方向([1])。

环形RNA(circRNA)是一种非编码RNA,最初被误解为Pre-mRNA错接的非功能产物。目前,circRNA已被证实可以调控多种分子的功能,包括非编码RNA、mRNA、DNA和蛋白质,从而调节细胞的生理和病理活动,越来越多的证据表明,circRNA在肿瘤的发生、发展以及对放疗和化疗的敏感性中起着至关重要的作用。放疗和化疗是大多数癌症的两种主要干预手段,但它们的治疗效果通常受到固有和获得性耐药的影响。因此,迫切需要开发新的策略来改善治疗反应。
circRNA的发生和功能
保守的circRNA的长度从几百个到几千个核苷酸不等。它们通常是由pre-mRNA转录子的非顺序反向剪接或线性RNA的反向融合产生的。具有3‘和5’端的circRNA共价连接形成环状结构,没有游离端,不受RNase R和外切酶降解的影响。这赋予了它们比线性转录本更高的稳定性。circRNA经典和主要功能是作为特定miRNA的分子海绵,调节mRNA的稳定性和翻译。此外,circRNA还作为蛋白质的海绵,作为蛋白质、mRNA和DNA的增强剂或协调剂,以及作为翻译的模板。
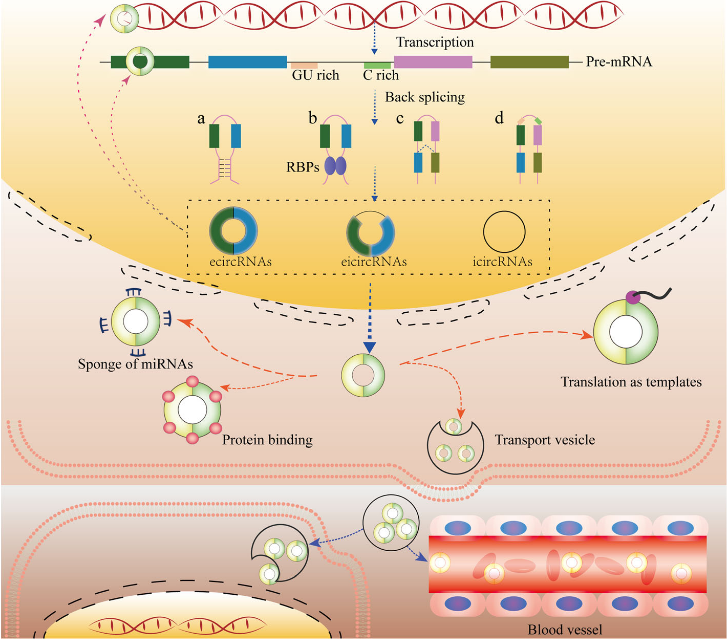
图1 circRNA的发生及功能([1])
circRNA在肿瘤耐药中的作用
circRNA是一种很有希望用于临床诊断和预后的肿瘤生物标志物,因为它们在体液中具有很高的稳定性和丰度。根据circRNA在癌症中的功能,有效的circRNA大致可分为两类:抗药抑制剂和抗药增强剂。
肿瘤耐药的分类
肿瘤细胞对放疗和化疗的反应。完全敏感的反应是理想的,但很少见,而放疗和化疗通常会受到后天或内在耐药的阻碍。获得性耐药大致可分为肿瘤干细胞(CSC)介导的获得性耐药和非CSC介导的获得性耐药。CSC介导的耐药归因于CSC的潜在增殖和分化能力,导致肿瘤复发或通过转移到新的位置种植新的肿瘤。非CSC介导的耐药因素包括治疗应激诱导的继发突变、肿瘤异质性、内质网应激改变、自噬、药物分布和代谢等。固有耐药机制与介导获得性耐药的机制相似,但不同的是固有耐药的特点是固有的,而不是由治疗压力诱导的。
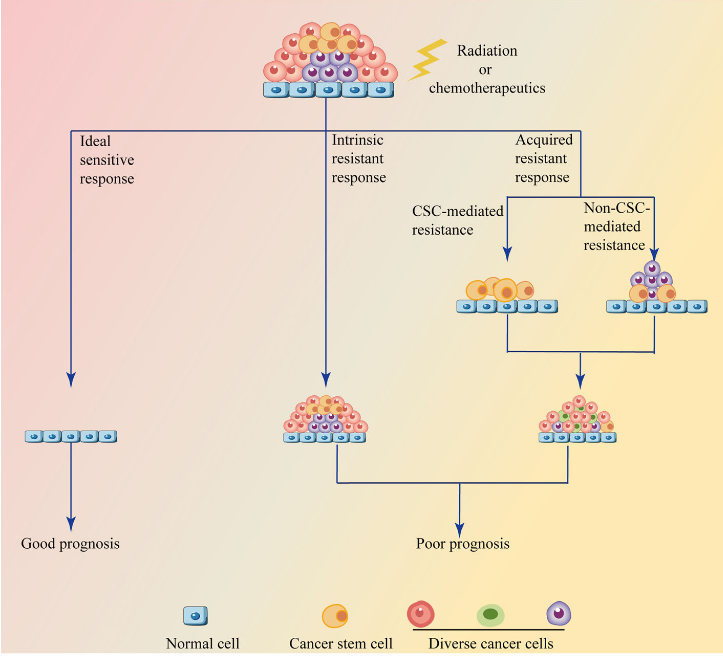
图2 肿瘤耐药的分类([1])
circRNA调控肿瘤细胞对放化疗反应的机制
药物通道和转运体的重构
药物浓度降低是产生化疗耐药性的主要原因。这可能是由于药物滞留在细胞内的囊泡和隔室,增加了药物外排或减少了药物影响。这些变化可能归因于药物通道和转运体的重构,包括ATP结合盒(ABC)蛋白家族、溶质载体和容量调节阴离子通道(VRAC)。ABC转运蛋白可以有效地将相应的底物(如化疗药物、激素和脂质)转运到胞外室、特定的细胞器和外泌体。通过这些过程,可以改变这些底物的吸收、代谢和活性。因此,ABC转运蛋白是逆转多药耐药(MDR)有希望的靶点。circSETD3可以与miR-520结合,增加ABCG2的水平,从而减少吉非替尼的细胞内蓄积,降低药物的敏感性。
血脑屏障(BBB)阻止了许多药物进入大脑,导致胶质瘤对化疗的反应不良,体外研究表明,circRNA DENND4C作为miR-577海绵,通过减少紧密连接相关蛋白的表达,增加了血脑屏障的通透性。这使得更多的DOX能够穿入体外模拟血脑屏障,导致胶质瘤细胞凋亡。
增加干细胞和上皮向间充质转化(EMT)
CIRS-7在骨肉瘤(OS)和食管鳞癌(ESCC)中表达上调。它与肿瘤进展呈正相关,并通过调节miR-7和miR-876-5p促进EMT。EMT是一种表观基因程序,通过它细胞失去了上皮表型,获得了间充质细胞的特性。EMT诱导转录因子,如twist和 snail,去促进ABC转运蛋白产生。它不仅增强了EMT,而且还上调了ABC转运蛋白的表达。沉默这些诱导EMT的circRNA成功地逆转了乳腺癌细胞中EMT相关的多药耐药。例如,circSMAD2能够通过降低miR-629的表达来抑制EMT,这是提高治疗敏感性的潜在靶点。
在大肠癌和胃癌中,circ_001680和circ-NOTCH_1分别升高。它们通过激活EMT或衰老诱导肿瘤干细胞(CSC)的产生是癌症治疗失败的另一个原因。CSC能够休眠很长一段时间以躲避有害的压力,如放疗和化疗。一旦有害的压力被消除,CSC可能在原发灶或转移灶分化和增殖,然后侵犯其他器官,导致肿瘤的复发或转移。
下游通路的调控
circATRNL1来源于ATRNL1基因,是口腔鳞状细胞癌(OSCC)中表达水平降低的一种失调的circRNA。当circATRNL1上调时,口腔鳞癌细胞的放射敏感性增强。这是因为它海绵miR-23a-3p以增加PTEN的转录,然后降低AKT的磷酸化。
食管鳞癌组织中circVRK1水平降低,且circVRK1水平低的患者预后较差。这可能是因为circVRK1作为miR-624-3p的海绵,降低了PTEN/PI3K介导的AKT活性,抑制了EMT,从而增加了食管癌细胞对放疗的敏感性。
自噬的重塑
自噬是一把双刃剑,温和的自噬有利于细胞在正常或轻度应激条件下存活,而过度的自噬会导致细胞死亡。治疗抗药性可能归因于自噬介导的氧化应激变化和自噬的关键代谢产物活力。circ_0009910在伊马替尼耐药的白血病细胞中表达增加。circ_0009910通过miR-34a-5p增强ULK1和触发自噬,从而促进了慢性粒细胞白血病(CML)的化学抗性。circEIF6在甲状腺乳头状癌(PTC)组织中高表达,并且负向调节miR-144-3p表达。将TPC1和BHT101细胞暴露于顺铂会进一步增加circEIF6并降低miR-144-3p表达。 circEIF6上调促进顺铂诱导的自噬。因此,它通过miR-144-3p /TGF-α轴增强了PTC细胞对顺铂的耐药性。
增强或削弱DNA修复能力
诱导DNA损伤是放化疗的另一个主要作用,通常会导致细胞死亡。然而,DNA修复会降低癌细胞对治疗的敏感性。BRCA1和BRCA2是DNA修复的两个关键酶,任一突变均会导致细胞对死亡刺激敏感,如顺铂、聚腺苷二磷酸核糖聚合酶(Parp)抑制剂和辐射。hsa_circ_0000199激活DNA修复分子his-tone家族成员X(H2AX)的潜力,导致对顺铂的耐药性增加。缺氧条件下,肝癌细胞中cZNF292的表达上调不依赖于缺氧诱导因子-1α。沉默cZNF292可以抑制辐射诱导的γ-H2AX病灶的形成,因为它与SOX9蛋白结合,从而抑制了β-catenin介导的DNA修复途径的活性,这最终增强了肝癌细胞对射线的敏感性。
肿瘤微环境的调控
TME是一个由微生物区系、酸性pH、炎症因子、基质金属蛋白酶、细胞外基质(ECM)、肿瘤相关成纤维细胞(CAF)、肿瘤相关巨噬细胞(TAM)等组成的复杂生态系统。原发肿瘤和相应转移瘤的TME甚至不同,这也影响癌细胞的药物敏感性。ciRS-7,已被证明调节TME的各种特性,它与好几种类型的癌症对化疗的敏感性相关。如,耐培美曲塞和顺铂的肺癌细胞中ciRS-7和circPVT1的表达水平升高。它们分别通过抑制EGFR/PI3K通路和miR-145-5p/多药耐药蛋白1(MRP1或ABCC1)轴来降低化疗(培美曲塞和顺铂)的效率。
参考文献
[1] Cui C, Yang J, Li X, Liu D, Fu L, Wang X. Functions and mechanisms of circular RNAs in cancer radiotherapy and chemotherapy resistance. Mol Cancer. 2020;19(1):58. Published 2020 Mar 14. doi:10.1186/s12943-020-01180-y