胖志
图片摘要:心血管疾病顶级杂志Circulation Research发表非编码RNA重要综述
2020年4月24日,德国法兰克福歌德大学的tefanie Dimmeler在Circulation Research(IF=15.862)杂志上发表了一篇题为“Noncoding RNAs in Vascular Diseases”的综述,对调节血管系统不同类型的非编码RNA进行介绍,包括miRNA、YRNA、snoRNA、circRNA。并重点介绍lncRNA在血管中的功能及研究面临的挑战([1])。

随着深度测序技术的出现,大量非编码RNA被挖掘,它们在心血管疾病中的作用也是逐渐被关注。miRNA的研究已经有近十年,已发现几种miRNA具有保护或者损伤血管的功能,并可以作为治疗靶点。现阶段,其它非编码RNA成为了研究的潮流,如环状RNA(circRNA)、YRNA或lncRNA。最初的研究表示它们可能在体内作为血管疾病的介质或拮抗剂。本篇综述文章简要介绍了控制血管系统不同类型的非编码RNA,随后着重介绍了lncRNA在血管中的作用。最后,指出lncRNA研究中面临的主要挑战。
MicroRNA
MicroRNA(miRNA)是约22个核苷酸的短非编码RNA(ncRNA),通过介导Argonaute(AGO)蛋白到达mRNA的3′非翻译区(UTR)的靶位点,阻断其翻译或诱导mRNA降解。许多miRNA参与血管生长,重塑和炎症的调节。例如,富含在内皮细胞(EC)的miR-126,可将其加工为miR-126-3p和miR-126-5p,两者在控制EC功能和减少动脉粥样硬化病变中都起着重要作用。相反,miR-92a会促进EC功能障碍,抑制血管生成,并促进动脉粥样硬化病变,并且针对miR-92a的拮抗剂已经投入到I期临床研究。炎症在动脉粥样硬化中起到关键作用,它也受miRNA的调节。例如,miR-155通过激活炎症和干扰巨噬细胞的脂质处理来增加动脉粥样硬化病变的形成。其他几种miRNA,例如miR-33,miR-122和miR-148a,可调节胆固醇的稳态和脂肪酸代谢,从而影响动脉粥样硬化病变的发生。最后,平滑肌细胞(SMC)受多种miRNA的控制,包括miR-143和miR-145,它们在血管损伤后会减少,它们的过表达会促进SMC的分化和抑制新内膜形成。
YRNA、 snoRNA和其他非编码RNA
不同于miRNA,关于其它短的和中等大小的RNA在心血管系统中的作用了解较少。例如,YRNA在人类中包含4个高度保守的转录本(RNY1、RNY3、RNY4和RNY5),大小在80至110个核苷酸之间,可以形成茎环结构。在心血管系统中,YRNA衍生的小RNA(s-RNY,长度约为24-34个核苷酸)作为一类与病变巨噬细胞凋亡相关的生物标志物和冠状动脉疾病风险的评估指标。此外,在培养的单核/巨噬细胞中,s-RNY可激活天冬氨酸蛋白酶依赖性细胞死亡和NF-κB依赖性炎症,细胞外s-RNY刺激受体细胞的死亡与Ro60相关。含有YRNA片段的细胞外囊泡通过调节IL-10赋予心脏保护作用。
snoRNA是核仁内的小非编码RNA,大小在60-170个核苷酸之间,可以指导其它RNA的化学修饰,尤其是在核糖体和小核RNA(snRNA)中发挥作用。将snoRNA分为两个不同的家族,box C / D家族,催化RNA甲基化,box H / ACA家族,催化假尿苷化。这两个家族都与一组特定的蛋白质结合形成snoRNP(小核仁核糖核蛋白),其中RNA提供靶向特异性。box C / D和H / ACA snoRNA的一个分支,称为scaRNA,定位于核Cajal体,在那里它们有助于剪接小体snRNA的修饰。越来越多的证据表明,snoRNA在心血管疾病中具有重要作用。例如,患有法洛氏四重症(最常见的先天性心脏缺陷)的婴儿左心室中有12种scaRNA减少,受影响的scaRNA仅靶向snRNA(剪接体的主要成分),导致受Wnt通路调控的转录物异常剪接。在压力状况下,snoRNA也会发挥作用。核糖体蛋白L13a基因座中的转录本U32a、U33和U35a的丢失足以赋予体外对脂毒性和氧化应激的抗性并防止体内氧化应激的传播。U17 snoRNA对HUMMR mRNA具有负调控作用,在U17 snoRNA缺陷细胞中,HUMMR的上调促进了内质网-线粒体的接触,降低了胆固醇的酯化,并促进了胆固醇向线粒体的运输。
增强子RNA和假基因的转录本在控制血管稳态或疾病中也可能具有重要功能。增强子相关的lncRNA LEENE促进RNA聚合酶II在eNOS(内皮型一氧化氮合酶)启动子上的募集,从而促进eNOS转录。最近发现一种缺氧控制的增强子RNA HERNA,它是由心脏的病理性应激诱导的,抑制HERNA可以保护小鼠免受应激诱导的病理性肥大。
circRNA
circRNA主要由编码蛋白质的外显子反向剪接产生的单链共价闭合环状RNA,因为没有5‘-帽和3’-poly(A)尾,对核酸外切酶有很高的抵抗力。cLrp6由低密度脂蛋白受体相关蛋白6 pre-mRNA的第84到87外显子环化形成,并作为miR-145的海绵促进体外血管SMC的增殖和去分化。抑制cLrp6可增加体内保护动脉粥样硬化的miR-145的表达并减少新内膜的形成。在动脉粥样硬化中,lncRNA ANRIL的外显子被转录生成circANRIL,circANRIL通过干扰核糖体RNA成熟来特异性调节SMC的死亡和增殖,从而对动脉粥样硬化起到保护作用。其他几种circRNA可以调节EC功能,包括cZNF292(环状锌指蛋白292)和cHIPK3(环状同源相互作用蛋白激酶3),前者是一种低氧诱导的circRNA,可控制EC增殖和血管生成,后者可诱导糖尿病患者视网膜血管异常。
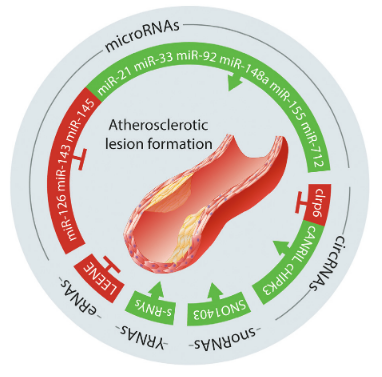
图1 一些非编码RNA调节血管疾病
lncRNA
MALAT1
lncRNA是长度大于200个核苷酸的非编码RNA,它在非编码RNA中占比最多,并且由很多不同的转录本组成。基因间非编码RNA MALAT1最早被发现作为肺癌转移的预后分子。MALAT1可以平衡EC的增殖和迁移,MALAT1的沉默增加了EC的迁移,同时抑制细胞增殖、产后视网膜血管生成和局部缺血引起的新血管生成。在糖尿病视网膜病变(一种涉及内皮细胞过度增殖的微血管疾病)显示MALAT1水平升高,沉默MALAT1可以减少视网膜血管损伤和炎症从而改善视网膜功能。与视网膜血管生成不同,MALAT1的沉默介导血管壁炎症反应的增加来促进动脉粥样硬化病变。此外,MALAT1沉默也增加了巨噬细胞的黏附、TNF(肿瘤坏死因子)和诱导型一氧化氮合酶的表达。在斑块中MALAT1表达水平的降低与较差的预后相关。
从机制上讲,竞争内源性RNA(ceRNA)机制,MALAT1通过抑制miRNA来控制基因表达。另一种是RNA-蛋白质互作的机制,例如,直接靶向VEGFR2(血管内皮生长因子受体2)促进血管生成。值得注意的是,MALAT1可以编码一个高度结构的短转录本mascRNA,它本身可能具有特定的功能。
MEG3
lncRNA MEG3在许多组织中都有高水平的表达,早期被认为是通过介导p53的积累来调节关键的抑癌基因。在体内,GapmeR介导的老年小鼠MEG3沉默促进了后肢缺血后新生血管的形成。在胚胎小鼠大脑中,MEG3的沉默导致血管生成通路的上调,包括VEGF(血管内皮生长因子)和Notch通路的基因,证实了MEG3对血管生成的负向调控。MEG3在平滑肌细胞中也存在作用,与其在内皮细胞中的抗增殖作用相反,MEG3被证明能诱导血管和肺动脉SMC增殖,这可能表明MEG3存在细胞类型特异性的活性。在缺氧性肺动脉高压中,MEG3上调并参与缺氧诱导的肺动脉平滑肌细胞增殖和细胞循环。还有证据表明MEG3介导炎症发挥作用,它的沉默可以阻止了oxLDL诱导的巨噬细胞的激活。
机制上,一种是假说是RNA-蛋白质的互作机制,例如MEG3通过与p53相互作用来抑制VEGF-A的转录。另一种是miRNA调控机制,MEG3被证明是miR-9的海绵,从而控制EC的增殖和血管生成。
SENCR
SENCR是血管细胞中富含的lncRNA,从FLI1基因的5‘端转录而来,以2种剪接突变体的形式存在。SENCR定位于细胞质,稳定平滑肌细胞的收缩表型,从而抑制迁移。体外高糖处理的SMC和糖尿病小鼠SMC中SENCR的表达水平均下调,SMC中SENCR的过表达逆转了高糖诱导的迁移效应。在内皮细胞中,SENCR可能通过调控迁移和血管生成基因,其中包括促血管生成基因CCL5和CX3CL1,来诱导人脐静脉内皮细胞(HUVEC)的增殖、迁移和血管生成。机制上,RNA pull down和质谱分析鉴定CKAP4是SENCR的相关蛋白,SENCR的沉默使CKAP4从细胞内转移到细胞表面。CKAP4与粘附连接蛋白CDH5结合,触发其内化,最终干扰膜的完整性,从而增加内皮通透性。
MANTIS
MANTIS是由ANXA4反义转录的内含子lncRNA,经表观遗传调控的lncRNA筛选鉴定。MANTIS的表达受组蛋白去甲基化酶JARID1B的控制,MANTIS的沉默会抑制血管生成。将人的EC移植到免疫缺陷小鼠体内,证实了MANTIS沉默的抗血管生成活性。机制上,鉴定出染色质重塑复合物SWI/SNF的解旋酶BRG1可以与MANTIS相互作用。这种互作是核小体重塑所必需的,通过诱导染色质重塑产生RNA聚合酶II更容易结合的结构,调节关键的内皮基因,包括SOX18,SMAD6和COUP-TFII。有趣的是,基于过表达的突变分析发现,lncRNA序列中的小Alu元件对于调控EC功能非常重要,因为这些序列足以抑制MANTIS靶基因ICAM-1(细胞间粘附分子1)。此外,观察到MANTIS沉默后ICAM-1转录起始位点上BRG1的增加,可以得出结论,MANTIS通过阻止BRG1结合来抑制ICAM-1转录。与这些发现一致的是,MANTIS也限制了ICAM-1介导的单核细胞粘附。综上所述,MANTIS在内皮细胞中起到动脉粥样硬化保护的作用。
PUNISHER
PUNISHER是由细胞阶段特异性RNA测序鉴定出来的lncRNA,在终末分化的EC中表达。PUNISHER的表达与血管发育的基因呈正相关,与细胞周期调节因子、染色质修饰调控分子和DNA损伤反应基因呈负相关。斑马鱼中PUNISHER功能丧失导致EC功能受损和血管损伤,包括分支和血管形成能力受损。回复实验中,使用人的PUNISHER恢复了部分血管功能。总之,PUNISHER的沉默使EC从有丝分裂和细胞分裂转向细胞粘附和细胞外相互作用模式。然而,并没有研究阐明PUNISHER潜在的分子机制。
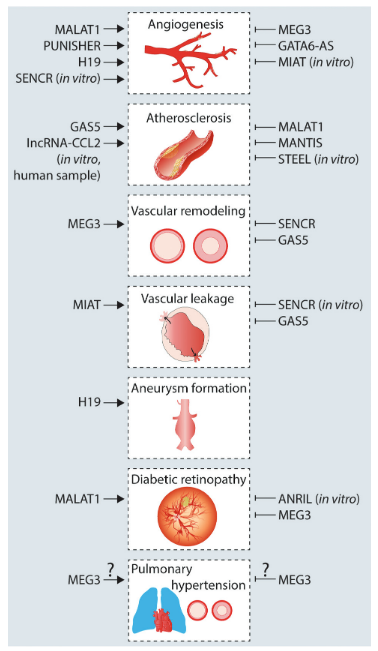
图2 lncRNA调节血管相关的疾病
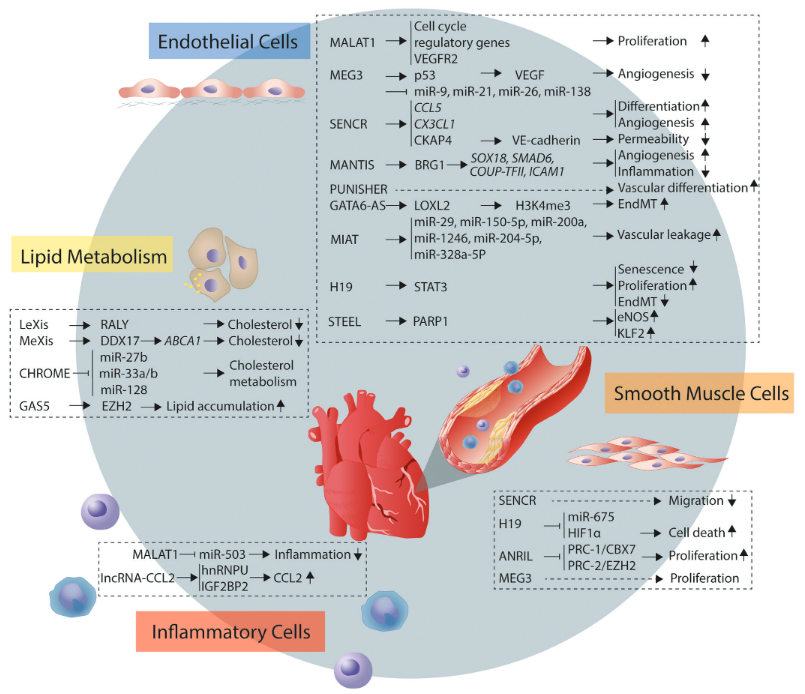
图3 lncRNA调控调控血管细胞功能的机制
lncRNA控制血管功能和脂质稳态
LeXis and MeXis
两个基因间的lncRNA LeXis和MeXis是细胞和系统胆固醇稳态的转录调节因子,可以调控LXR(肝脏X受体)。LXR激活诱导胆固醇外排相关基因的表达,通过介导脂肪酸合成促进胆固醇酯化,并抑制低密度脂蛋白受体装载胆固醇,这一效应是通过LeXis与异质核蛋白RALY的相互作用介导的。MeXis,可以充当Abca1基因LXR依赖性转录的扩增子,该基因对于调节胆固醇的外流至关重要。MeXis与转录辅助因子DDX17相互作用并引导其与启动子结合,从而导致细胞对胆固醇超载的反应减弱,并加速动脉粥样硬化的产生。
CHROME
灵长类特异性lncRNA CHROME是细胞和系统胆固醇稳态的重要调控分子。用富含胆固醇的饮食喂养的非人类灵长类动物,在肝脏中显示出升高的CHROME表达水平。CHROME的过表达与在人类肝细胞和巨噬细胞中LXR激活诱导的胆固醇外流一致,表明CHROME在LXR的下游参与调控。机制上,细胞中CHROME的沉默降低了胆固醇外流和Abca1 mRNA的水平,但未降低Abca1 pre-mRNA的水平,表明调控发生在转录后水平。此外,CHROME降低miR-27b,miR-33a,miR-33b和miR-128的抑制活性,它们可抑制介导胆固醇转运的基因,从而鉴定出lncRNA的胆固醇调控网络。
STEEL
STEEL是由HOXD1-HOXD3基因座的反义链表达的EC富集的lncRNA。STEEL可以促进HUVEC在体外的网络形成和迁移,以及体内血管的形成。随后的功能研究确定STEEL是EC中基因表达的有效调节剂,主要影响反式基因的表达。有趣的是,STEEL促进了eNOS(血管舒张调节因子的调节剂)和KLF2(血液动力的重要传感器)的上调,并受它们反馈抑制。机制上,STEEL
介导PARP1复合物的形成,其中RNA有助于募集PARP1靶向色氨酸区域,例如KLF2和eNOS的启动子。
lncRNA在鉴定和研究中的主要挑战
- 与蛋白质编码的转录本相比,lncRNA的表达较低,这使最小阈值选择和单细胞测序方法较难。(b)lncRNA存在聚(A)尾巴的异质性,在制备文库时需要考虑。(c)lncRNA存在组织和细胞类型特异性表达。在这种情况下,有必要提及许多lncRNA的注释不足,并且在不同的组织中表达和功能有所不同。(d)lncRNA拷贝数的确定。在这方面,数字PCR可以对目标核酸进行绝对定量。(e)实验设计和结果分析。lncRNA经常定位于相邻基因或其它调控序列,因此,使用CRISPR可能对预期的lncRNA和周围基因产生影响。(f)lncRNA的保守性。与mRNA不同,lncRNA几乎没有同源线性转录物,但是lncRNA基因座可能在不同的物种之间是保守的,表明转录本可以在序列或者功能方面保守。(g)通常采用沉默的方法来验证lncRNA的功能,如果采用过表达的方法,产生具有多个剪接突变体的长转录本,技术上通常会面临挑战。
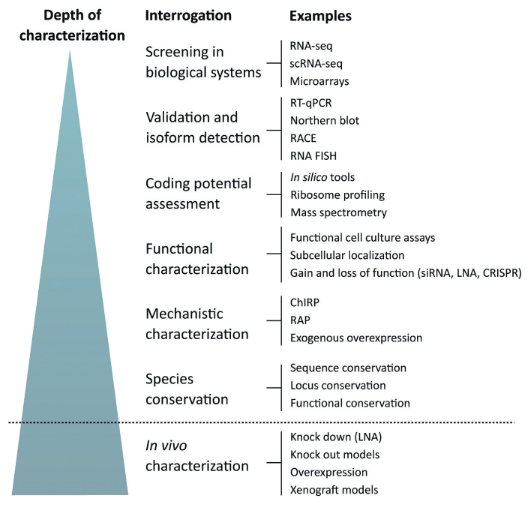
图4 lncRNA研究策略
参考文献 [1] Jaé N, Dimmeler S. Noncoding RNAs in Vascular Diseases. Circ Res. 2020 Apr 24 ;126(9):1127-1145.doi:10.1161/CIRCRESAHA.119.315938.