CDR1as是最早报道功能机制的circRNA分子,也是目前唯一实现CNS大满贯的circRNA分子。1月13日,Cancer Cell杂志报道了CDR1as在黑色素瘤中的功能机制研究,表明黑色素瘤中CDR1as是由LINC00632基因的转录产物加工形成的,可以被EZH2/PRC2沉默表达。CDR1as可结合IGF2BP3,CDR1as表达降低后可释放IGF2BP3,改变一些靶基因的表达状态,参与黑色素瘤的迁移侵袭及GPX4药物敏感性([1])。
CDR1as研究的主要进展:
- 2013年,两篇Nature报道CDR1as可竞争性结合miR-7,最早证明了circRNA具有miRNA Sponge的功能([2, 3])。miRNA Sponge功能模型也成为目前报道最多的circRNA功能模型。
- 2017年,Sicence报道敲除CDR1as小鼠大脑神经突触间信号传递异常([4])(推荐阅读:重磅!Science在线发表circRNA研究论文)
- 2018年,Cell文章报道人类大脑中RNA间相互作用的网络,报道了四种非编码RNA“相生相克”的精彩故事([5])
- 2019年,Cell报道心脏翻译组学研究,其中发现CDR1as可以结合在核糖体测序中被捕获,提示该分子也可能被翻译([6])。(推荐阅读:Cell:心脏翻译组学文章,证明circRNA可翻译)
- 截至目前,CDR1as的研究论文和综述已经多达85篇,是研究报道最多的circRNA分子。
在本文中,作者发现黑色素瘤中CDR1as表达沉默与肿瘤的浸润侵袭状态有关,因此深入分析了黑色素瘤中CDR1as生成,表达,调控,功能方面的机制。作者发现CDR1as的表达与上游非编码RNA基因LINC00632密切相关,并证实了CDR1as的表达是受到LINC00632表达变化而影响的。随着黑色素瘤的进展,CDR1as表达会呈现下降趋势,EZH2/PRC2介导了LINC00632/CDR1as Locus的表达沉默。在黑色素瘤中miR-7不是CDR1as影响侵袭迁移能力的原因,分析表明IGF2BP3的结合与这些现象有关。本文从RNA结合蛋白入手分析了CDR1as 与黑色素瘤侵袭能力的关系,并基于多种在线数据和信息分析了相关的基因和通路,药物敏感性等。下面就让我们一起学习一下本文:(声明:为了清楚的表达文章的故事,有些内容与原文的顺序有所区别,敬请谅解)
CDR1as在黑色素瘤中的功能是怎样的?
作者分析了4例黑色素细胞和10例短培养黑色素瘤细胞(melanoma short-term cultures,STCs)的RNA-seq数据,分析了其中的circRNA表达情况。从测序数据中,作者分析了环状RNA与来源基因的线性RNA表达的比值,发现CDR1as的成环比例比预期的要低。10例STCs中6例检测不到CDR1as,作者发现另外的4例是从黑色素瘤脑转移的病灶分离的,怀疑是由于分离的标本中掺杂了胶质细胞的缘故。黑色素瘤的细胞系中CDR1as的表达也不高。进一步分析表明相对于初发的患者,高转移侵袭的黑色素瘤标本中CDR1as的表达显著降低。生存分析表明,CDR1as表达最低的25%患者生存情况明显比其余75%的患者要差。综合这些现象,CDR1as在黑色素瘤中表达如何调控的,它的功能和机制等问题就非常值得深入探讨。
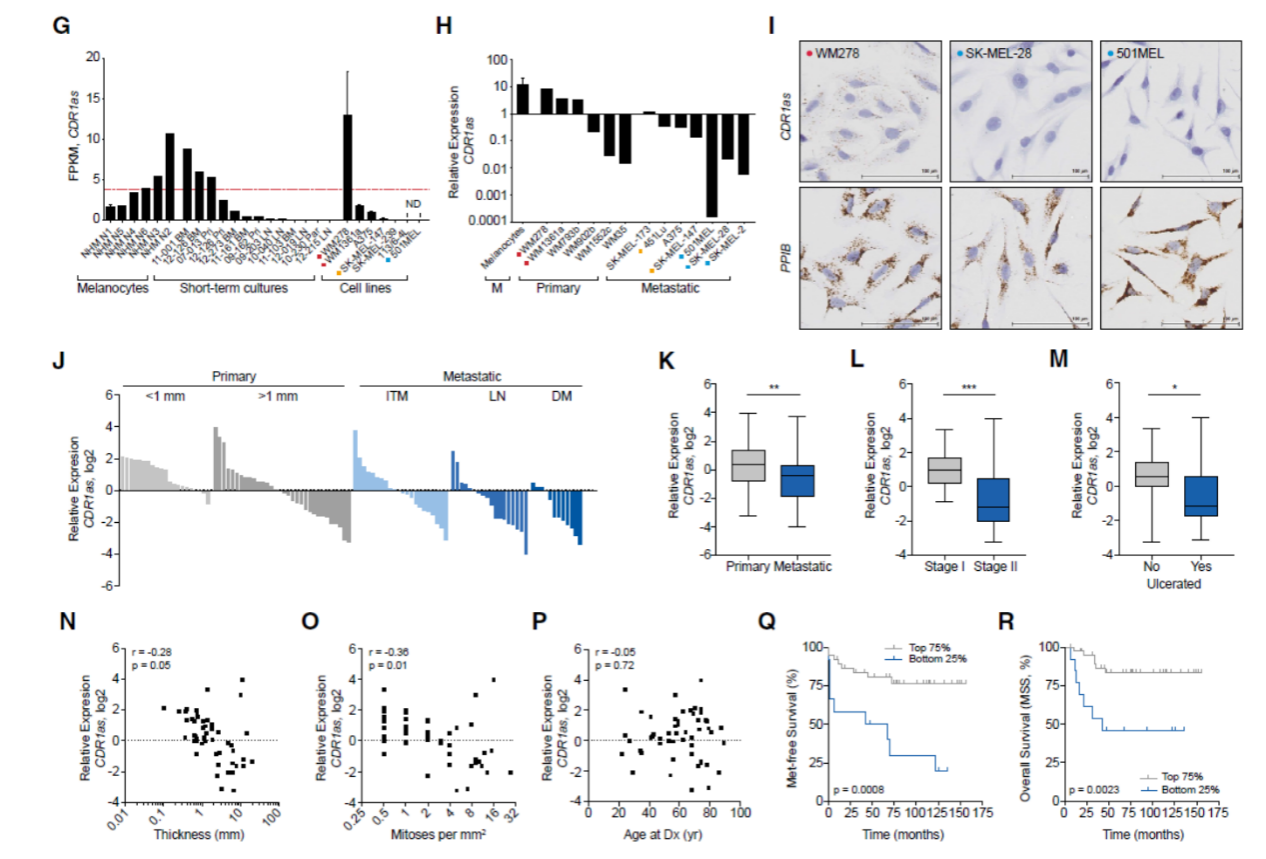
图1 黑色素瘤中CDR1as表达下调,与肿瘤侵袭有关 ([1])
为进一步证明CDR1as在黑色素瘤迁移侵袭中的作用,作者构建了Dox诱导表达靶向CDR1as的shRNA分子的模型,结果显示在低侵袭性高表达CDR1as的黑色素瘤细胞系(WM278和WM115)中干扰CDR1as并不能显著影响细胞增殖,但可以显著提高细胞侵袭能力。在低表达LINC00632/CDR1as的细胞(SK-MEL-28 和 501MEL)中,干扰CDR1as对侵袭表型没有影响。SK-MEL-28 和 501MEL细胞中过表达CDR1as能降低侵袭性,体内成瘤实验表明,干扰CDR1as并不显著改变肿瘤大小和生长速度,但可以显著体提高肺转移比例。
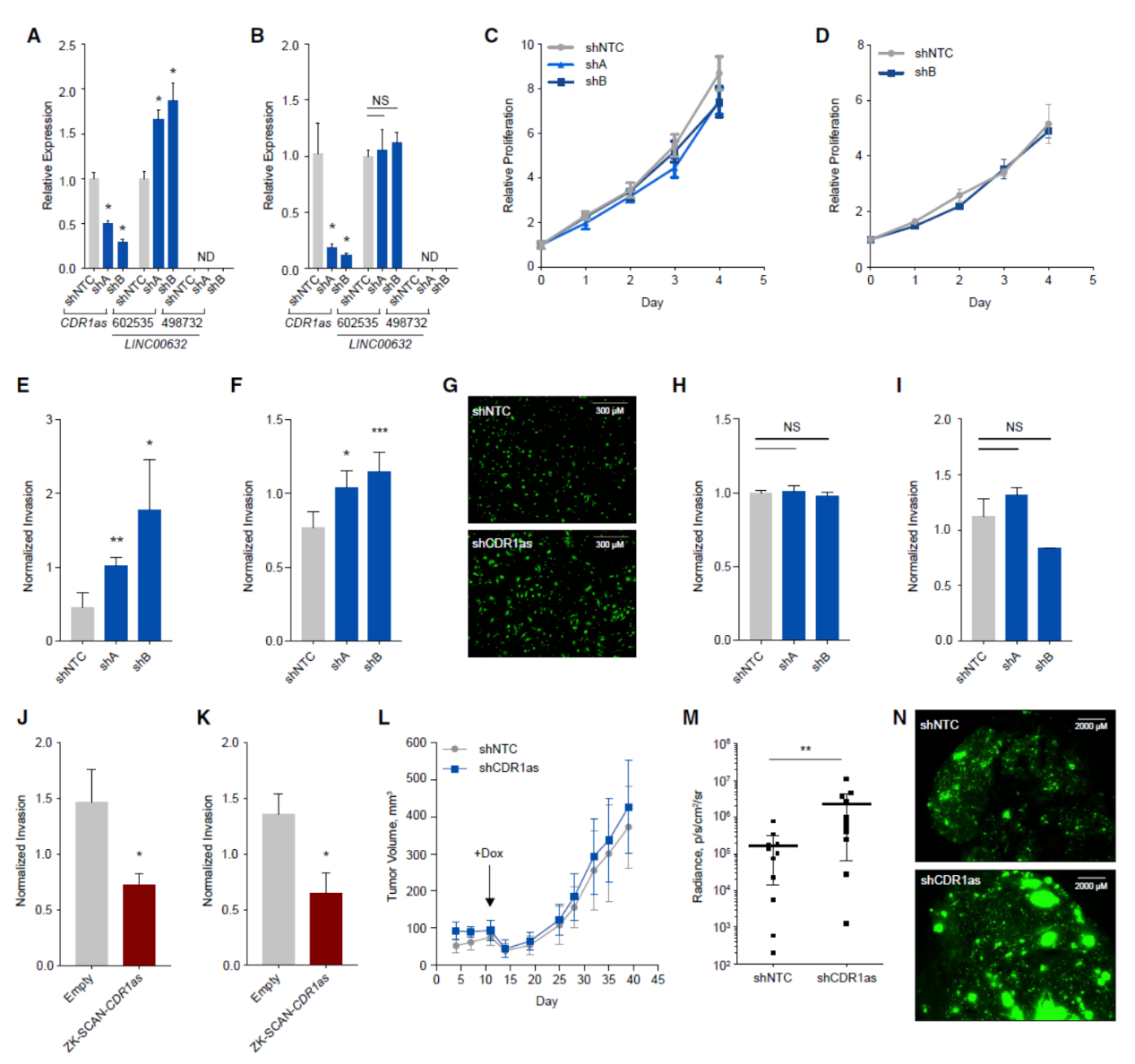
图2 CDR1as降低可促进黑色素瘤转移侵袭能力 ([1])
黑色素瘤中CDR1as的生成是如何调控的?
早期曾有报道发现CDR1as是在一个非编码RNA LINC00632的基因内部。本文作者进一步验证了这个假设,并证明CDR1as的表达的确与LINC00632高度相关。多种在线数据的统计结果均表明CDR1as的表达与LINC00632高度相关(相关系数r=0.86, p<0.0001),这些数据包括TCGA,GTEx,mouse neural crest-derived tissues。作者还从测序数据和PCR验证实验找到了LINC00632来源的两个转录本(ENST00000498732和ENST00000602535)包含了CDR1as序列,证明了CDR1as从LINC00632的产物中形成。Cas9破坏3’剪切位点可显著减少CDR1as的生成比例,但同时增加了 LINC00632的生成比例。SAM CRISPR/Cas9技术增强LINC00632的表达也会同步提高CDR1as的数目。这些数据均佐证了之前报道的CDR1as来源于非编码RNA基因LINC00632。
miR-671-5p可通过RNAi机制促进CDR1as的降解,但还没有报道表明LINC00632是miR-671-5p的作用底物,因此作者分析了黑色素瘤组织和细胞中三者的表达关系,结果表明在病人组织中miR-671-5p的表达与CDR1as的表达仅有微弱的相关性,但细胞中的表达没有相关性。过表达miR-671-5p可降低CDR1as的表达,但不影响LINC00632的表达,而敲降miR-671-5p并不能使得CDR1as表达值回复。综上现象,黑色素瘤中CDR1as的表达似乎并非由miR-671-5p的调控实现,更可能是由于CDR1as的来源基因LINC00632表达的影响造成的。
在线数据分析表明黑色素瘤中LINC00632基因区域(包括CDR1as在内)并不存在典型的删除突变,但H3K27me3和H3K27ac的ChIP结果强烈提示LINC00632基因区域在黑色素瘤中存在较强的染色体关闭信号。EZH2 抑制剂GSK126处理后可明显恢复LINC00632和CDR1as的表达。这表明黑色素瘤中LINC00632/CDR1as的表达是受到EZH2/PRC2的调控。
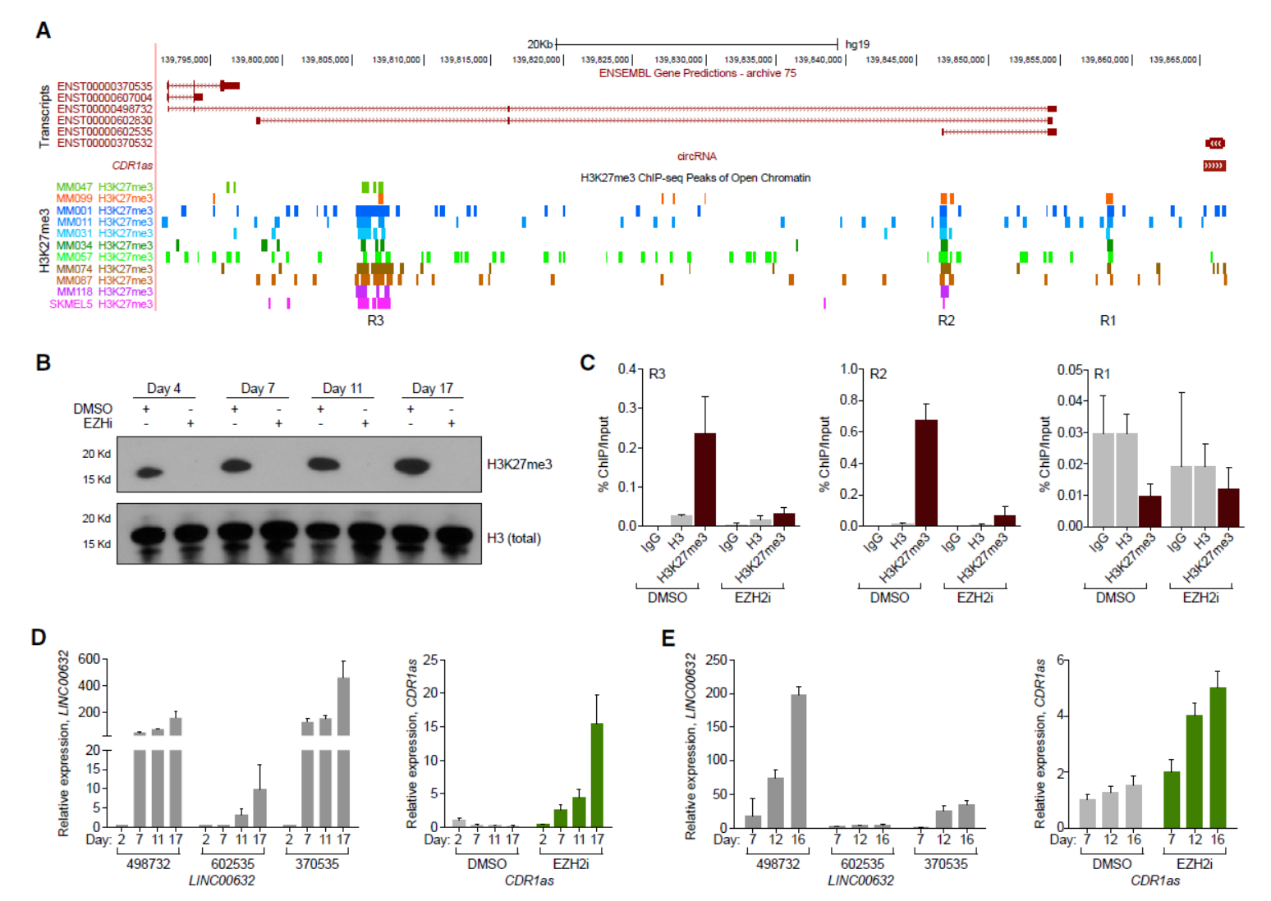
图3 LINC00632/CDR1as的表达是受到EZH2/PRC2的调控 ([1])
如何探讨黑色素瘤中CDR1as与miR-7的关系?
CDR1as是最早报道功能机制的circRNA分子,竞争性结合miR-7是最有代表性的miRNA sponge模型([2,3]),但CDR1as敲除的模型中也显示,在脑组织中CDR1as还具有稳定miR-7的作用([5])。在黑色素瘤中CDR1as与miR-7的关系是怎样的呢?
干扰CDR1as并不影响miR-7-5p的表达,也不会对miR-7结合位点荧光素酶报告基因产生影响,一些miR-7的下游基因也没有受到影响。Cas-9敲除miR-7-1可显著抑制miR-7表达,但不会改变细胞侵袭能力,反而在敲除miR-7-1的基础上干扰CDR1as可提高细胞侵袭能力。这说明黑色素瘤中敲降CDR1as导致的侵袭能力增强并非由miR-7介导。
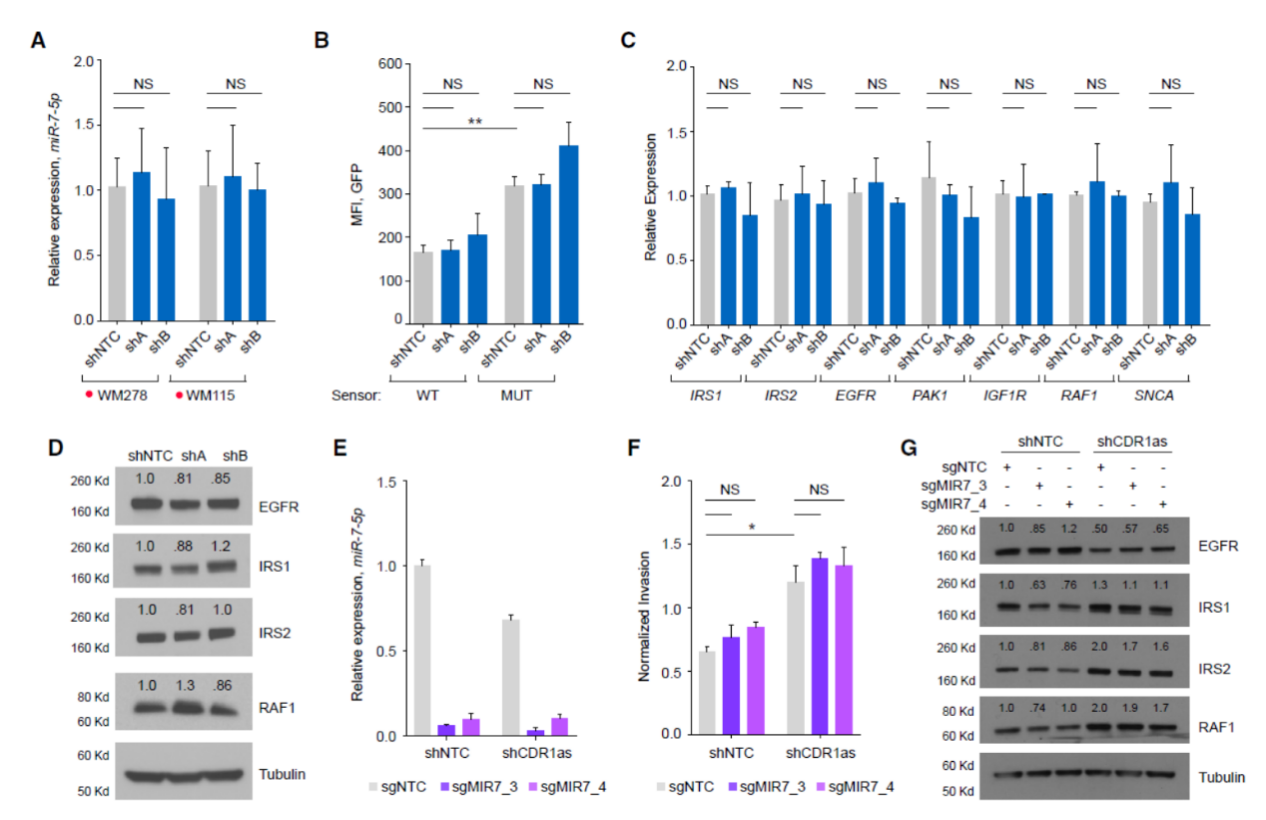
图 4 黑色素瘤中敲降CDR1as导致的侵袭能力增强并非由miR-7介导 ([1])
黑色素瘤中CDR1as与IGF2BP3结合的作用机制
为揭示CDR1as在黑色素瘤中的作用机制,作者首先思考了CDR1as可能的相互作用蛋白。与传统思路不同,作者首先从CLIP-seq的在线数据中分析了可能的结合蛋白,并从中发现了IGF2BP家族蛋白存在与CDR1as相互作用的证据。为进一步验证,作者进行了RIP分析,发现IGF2BP3对CDR1as的富集作用最强,并且对LINC00632的结合不高,说明IGF2BP3具有特异性结合CDR1as的作用。干扰IGF2BP3可降低由于干扰CDR1as导致的侵袭增强现象。IGF2BP3的RIP-seq能很强的富集CDR1as,作者在黑色素瘤中的数据与已发表的IGF2BP3的RIP-seq或CLIP-seq数据有20~40%的重叠。CDR1as序列中存在大量的IGF2BP3结合位点。
CDR1as干扰前后IGF2BP3结合的靶分子种类没有太明显的改变,但CDR1as干扰后变化显著的基因更倾向于富集到IGF2BP3结合的靶分子列表中。为进一步分析这些基因对CDR1as敲降后促进侵袭的表型的关系,作者进行了小规模RNA干扰文库的分析,针对CDR1as干扰后表达会增高的9种基因进行RNAi文库分析,其中6 种基因能显著逆转因干扰CDR1as导致的侵袭增加。其中SNAI2 和MEF2C在CDR1as干扰后的表达增高是由IGF2BP3介导的。
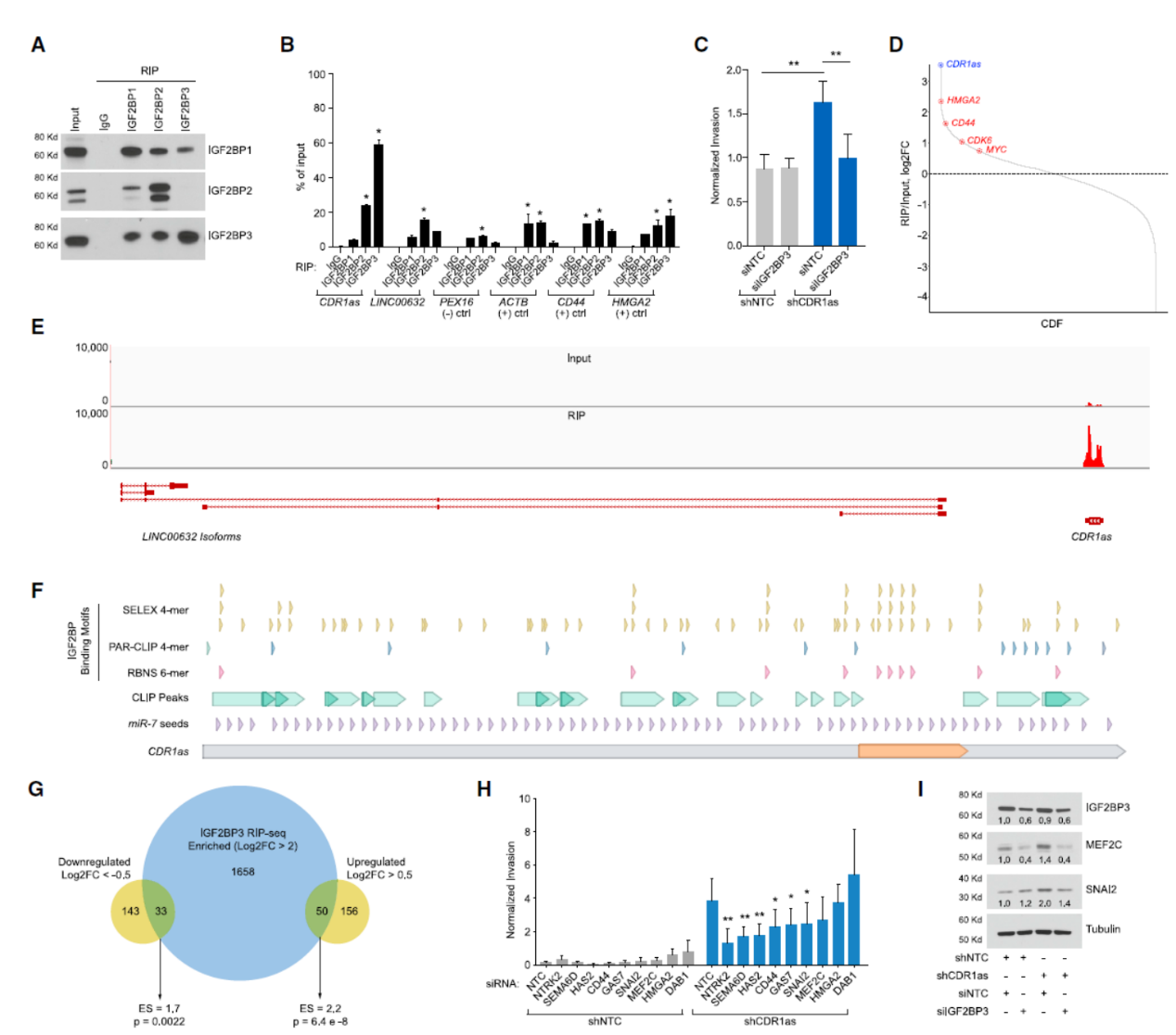
图5 IGF2BP3结合CDR1as,与CDR1as干扰后促进侵袭的表型有关 ([1])
最后,为分析CDR1as表达状态与黑色素瘤中药物敏感性和基因型的相关性,作者在Depmap和CTRP的数据中进行了挖掘,并结合CCLE的数据对所有相关细胞中CDR1as表达状态进行了分组,分为高表达,中表达和低表达三组。有趣的是,作者发现低表达CDR1as 组的细胞对多种MAPK通路的抑制剂更敏感。但在CDR1as高表达的细胞中敲低CDR1as并不能增加细胞对BRAF抑制剂(Dabrafenib)的敏感性。CDR1as表达值与BRAF突变的关系分析表明CDR1as低表达的细胞更高比例携带了BRAF突变。有趣的是,高表达CDR1as的细胞对GPX4抑制剂更敏感。sgRNA文库数据分析也表明,GPX4在CDR1as高表达的细胞系中富集显著高于CDR1as低表达组。黑色素瘤中GPX4抑制剂的敏感性与MITF / AXL基因的表达有关,MITF低表达且AXL高表达的细胞对GPX4抑制剂更敏感,但对MAPK抑制剂等药物不敏感。作者分析发现CDR1as高表达的细胞倾向于表现出MITF低表达和AXL高表达,而CDR1as低表达的细胞倾向于表现出MITF高表达和AXL低表达。实验结果也表明CDR1as表达状态与GPX4抑制剂敏感性有相关性,低表达CDR1as组的细胞对GPX4抑制剂更敏感。然而在高表达CDR1as的细胞中干扰CDR1as后并不能提高对GPX4的敏感性,说明CDR1as可作为黑色素瘤对GPX4抑制剂敏感性的指标,但并非该药物发挥功能的关键因素。 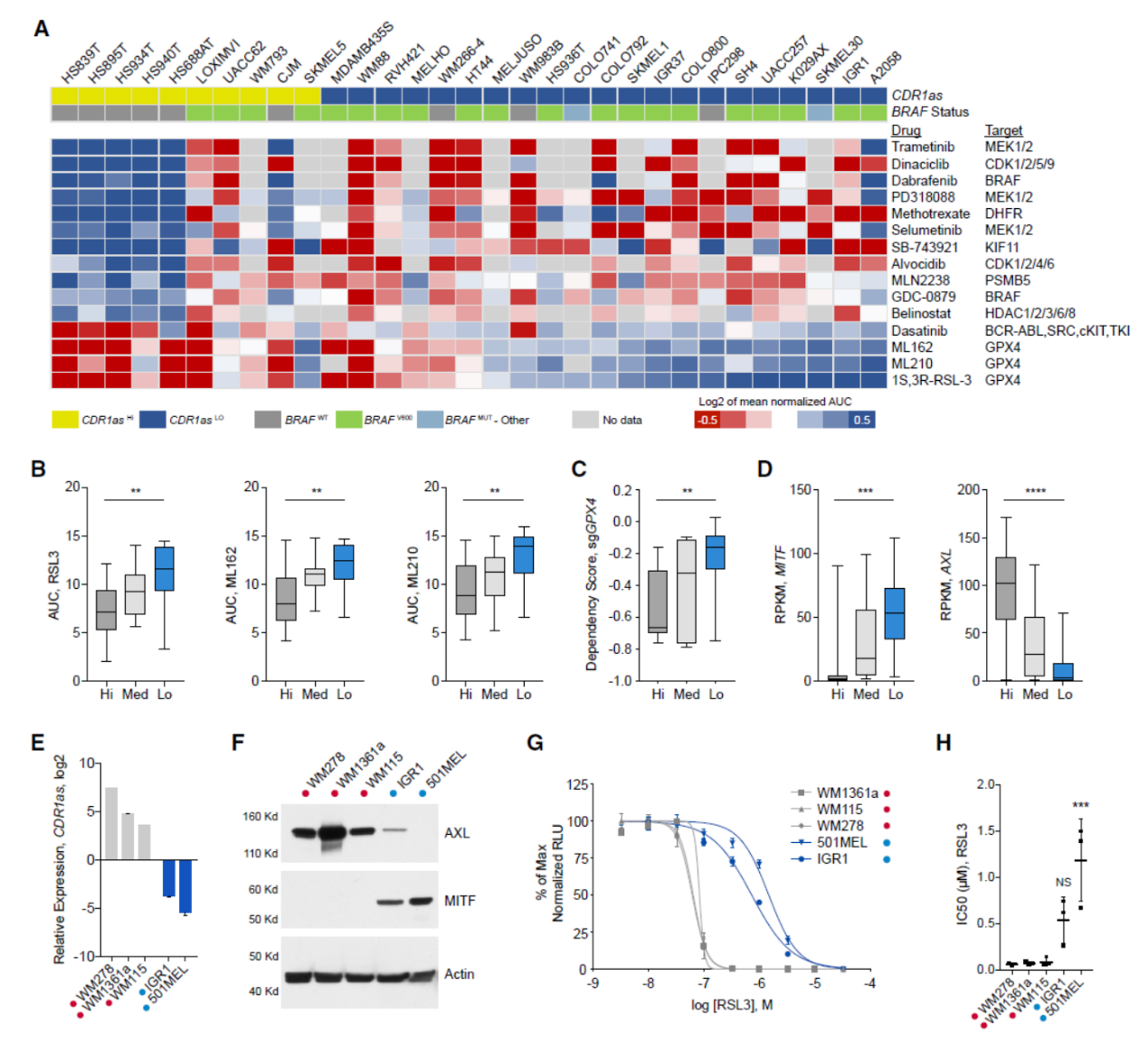
图6 CDR1as表达对应于MITF / AXL表达状态,与GPX4抑制剂敏感性有关 ([1])
本文的发现报道了CDR1as通过结合蛋白调控重要生理活动的机制,大大提高了对CDR1as的功能机制的认知。文章有很多值得学习的技术和思路,概括而言包括以下几点:
(1)在线资源的有效整合可帮助预测分析和设计相关实验。文中多次运用了在线数据资源,包括TCGA,GTEx,CLIP-seq,ChIP-seq以及后面分析药物敏感性和基因突变信息中用到的Depmap,CTRP的数据以及CCLE的数据。这些在线数据能大大提高研究思路和方向设计的效率,从已有数据中挖掘可能的机制和方向。这种做法非常值得学习,也越来越多的用到。
(2)RNA结合蛋白是突破circRNA研究机制单一性的重要思路。CDR1as是最早也是最有影响力的miRNA Sponge模型分子。但本文的实验分析过程中却发现miR-7对作者分析的模型并没有作用,因此提示存在其他的作用机制。生物体本来就是复杂和多维度的,从不同角度发现和认识生物分子的功能是非常必要的。
(3)高通量文库在探索功能性分子中的价值。本文并没有从一开始就利用高通量文库进行筛选分析,但在后面探索CDR1as下游基因与侵袭迁移表型及IGF2BP3结合作用之间的关系时用到了一个只有几个分子的RNAi文库体系,用于评价这些基因在相关通路中的作用。后面的Depmap数据则是高通量的RNAi和sgRNA文库体系的结果,间接的为本文最终的结论提供了帮助。
参考文献
1. Douglas H., et al., Epigenetic Silencing of CDR1as Drives IGF2BP3-Mediated Melanoma Invasion and Metastasis. Cancer Cell, 2020. 37(1): p. 55-70.
2. Memczak, S., et al., Circular RNAs are a large class of animal RNAs with regulatory potency. Nature, 2013. 495(7441): p. 333-8.
3. Hansen, T.B., et al., Natural RNA circles function as efficient microRNA sponges. Nature, 2013. 495(7441): p. 384-8.
4. Piwecka, M., et al., Loss of a mammalian circular RNA locus causes miRNA deregulation and affects brain function. Science, 2017. 357(6357).
5. Kleaveland, B., et al., A Network of Noncoding Regulatory RNAs Acts in the Mammalian Brain. Cell, 2018. 174(2): p. 350-362 e17.
6. van Heesch, S., et al., The Translational Landscape of the Human Heart. Cell, 2019. 178(1): p. 242-260 e29.