10月27日,Nature Communications杂志在线发表了台湾中央研究院Kuo Hung-Chih为通讯作者的文章,报道发现circBIRC6参与干细胞多能性调控[1]。

文章报道发现干细胞中特异的剪切因子ESRP1调控了circBIRC6的形成过程,circBIRC6通过竞争性结合miR-34a和miR-145参与调控干细胞多能性。ESRP1受到Oct4,Nanog等转录因子的调控。本文首次报道干细胞多能性相关的circRNA分子,是circRNA功能研究的重要扩展。那么作者是怎样确定circBIRC6与干细胞多能性的关系的?如何确定ESRP1调控circBIRC6的形成过程的?带着这些疑问,我们一起来拜读一下这篇文章吧:
作者如何筛选干细胞多能性相关circRNA 分子的?
作者筛选干细胞多能性相关的circRNA总体思路是:基于Nikolaus Rajewsky教授2013年发表在Nature的文章的数据,从中找出Reads数大于40的circRNA分子(共61种),然后基于RNase R消化后PCR验证,再分析hESC分化前后是否有特异性变化等,最终找到目标circRNA分子。
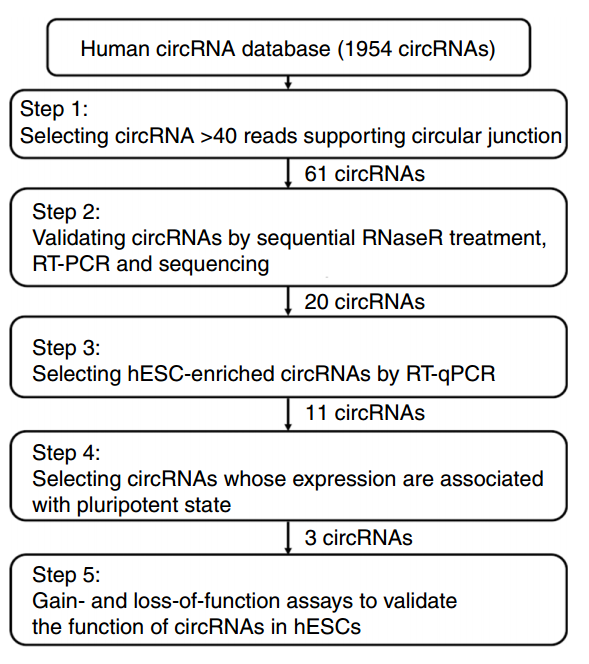
图1 干细胞多能性相关circRNA分子筛选方案流程(来自[1])
hESC分化的实验选择了体外分化胚状体(EB球)的体系,主要比较分化为EB球前后circRNA的表达变化,以及所对应的线性mRNA的变化情况。从数据库中初步筛选得到的61种circRNA分子,有21种可在hESC细胞中鉴定到,与分化为EB球后相比,有11种为hESC特异性表达的,其中9种是在分化为EB球后明显下调的。作者也在诱导多能干细胞(iPSC)体系中进行了验证,所有11种circRNA分子均在iPSC中富集。circRNA的表达经常会伴随对应线性基因的表达而变化,经过比对分析,作者发现三种circRNA分子的表达不与所对应的mRNA表达变化一致:circBIRC6、circMAN1A2及circILKAP。进一步,作者通过Northern杂交分析了circRNA表达量与干细胞多能性状态的相关性,表明有三种circRNA的表达呈现出强相关性,它们是:circBIRC6、circCORO1C和circMAN1A2。
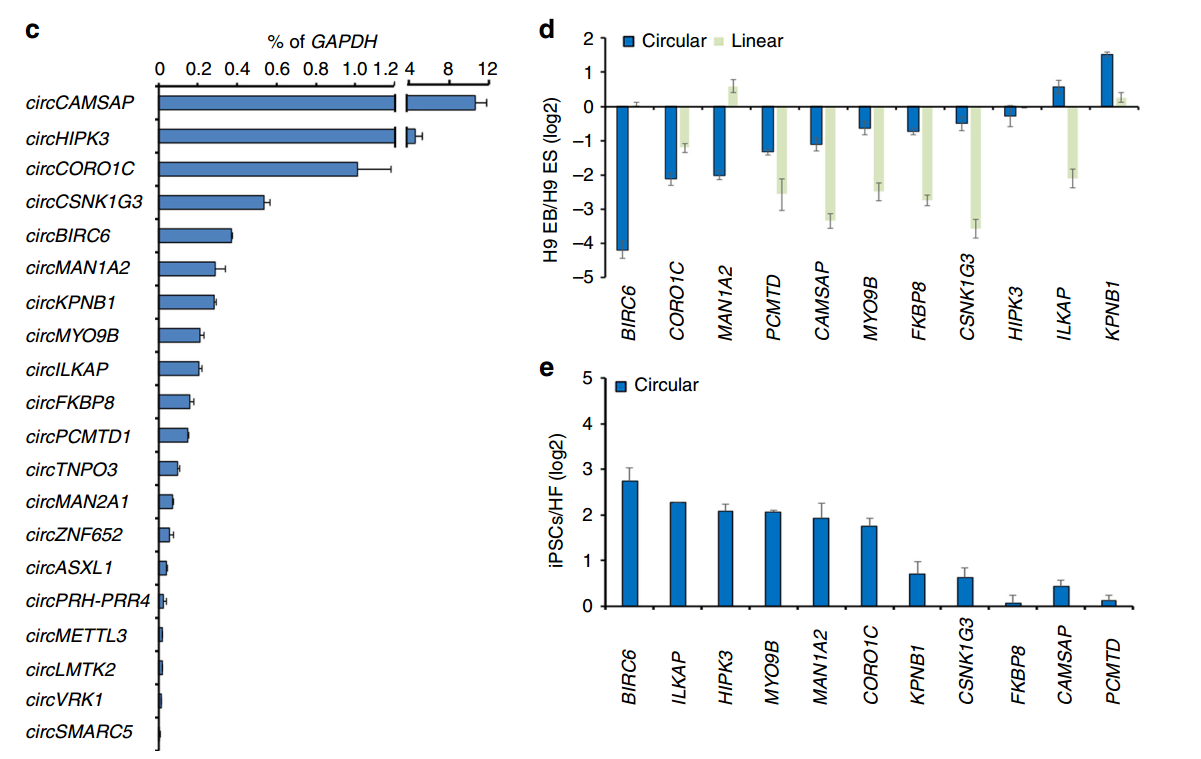
图2 体外分化实验分析多能性相关circRNA分子 (来自[1])
以上的实验说明了3种circRNA分子的表达状态与细胞的多能性状态的相关性,还不能说明这些circRNA分子参与了干细胞多能性的调控,还需要针对这些circRNA分子进行Knock Down分析。于是作者设计了RNA干扰实验进行分析,结果表明干扰circBIRC6和circCORO1C后AP染色阳性率明显减少,而干扰circMAN1A2后作用并不明显。QPCR和免疫荧光等实验分别分析干性及分化状态相关的转录因子或marker基因都表明干扰circBIRC6和circCORO1C后干细胞多能性状态明显受到影响,倾向于分化状态。干扰circMAN1A2后作用不明显。

图3 circBIRC6和circCORO1C与干细胞多能性相关 (来自[1])
circRNA是由mRNA的前体分子加工形成的,会不会这种前体分子也参与了这一作用过程?作者分别设计了专门针对线性RNA分子以及共同外显子区的shRNA分子,靶向相应外显子区的可同时干扰线性RNA和circRNA,而专门靶向线性RNA区域的shRNA则不会影响circRNA的含量。AP染色结果表明单独干扰BIRC6和CORO1C的线性RNA分子形式并不会影响细胞的多能性状态。QPCR和免疫荧光等实验分析多能性和分化相关的基因,结果表明是circRNA分子参与调控了细胞的多能性,多对应的线性mRNA并不会直接影响细胞的多能性状态。
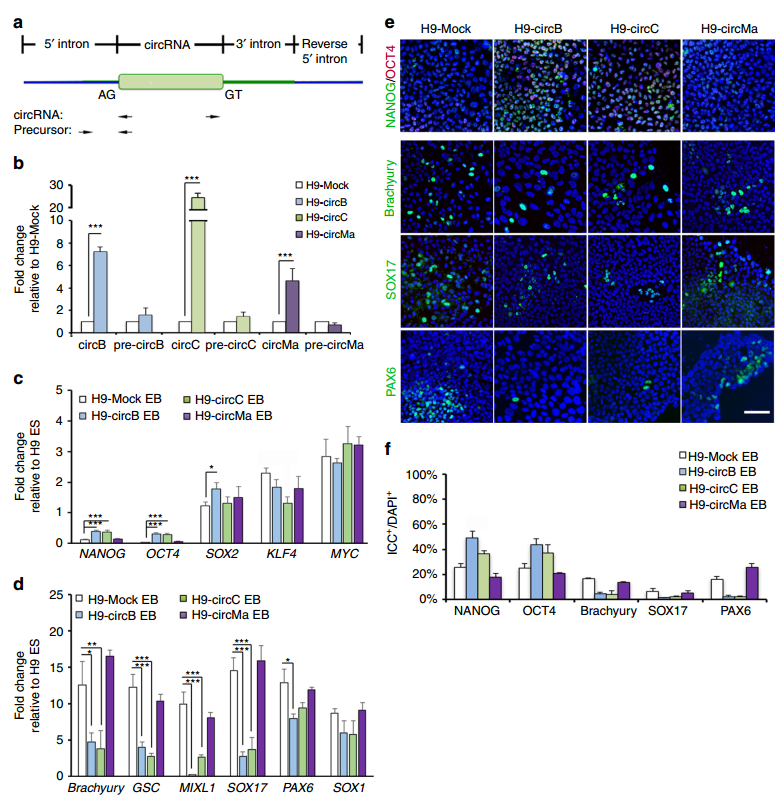
图4 circRNA而非线性RNA参与干细胞多能性调控 (来自[1])
干扰circBIRC6和circCORO1C可影响干细胞多能性状态,那过表达它们能否稳定或促进细胞转变为多能性的状态呢?会不会促进体细胞重编程?在hESC中过表达它们并不影响细胞的多能性状态。报告基因实验表明单独表达这些circRNA分子并不足以诱导细胞发生重编程。但在经典的四因子(Oct4、SOX2、Klf4和Myc,OSKM)诱导iPSC体系中,过表达circBIRC6和circCORO1C可明显促进细胞重编程。
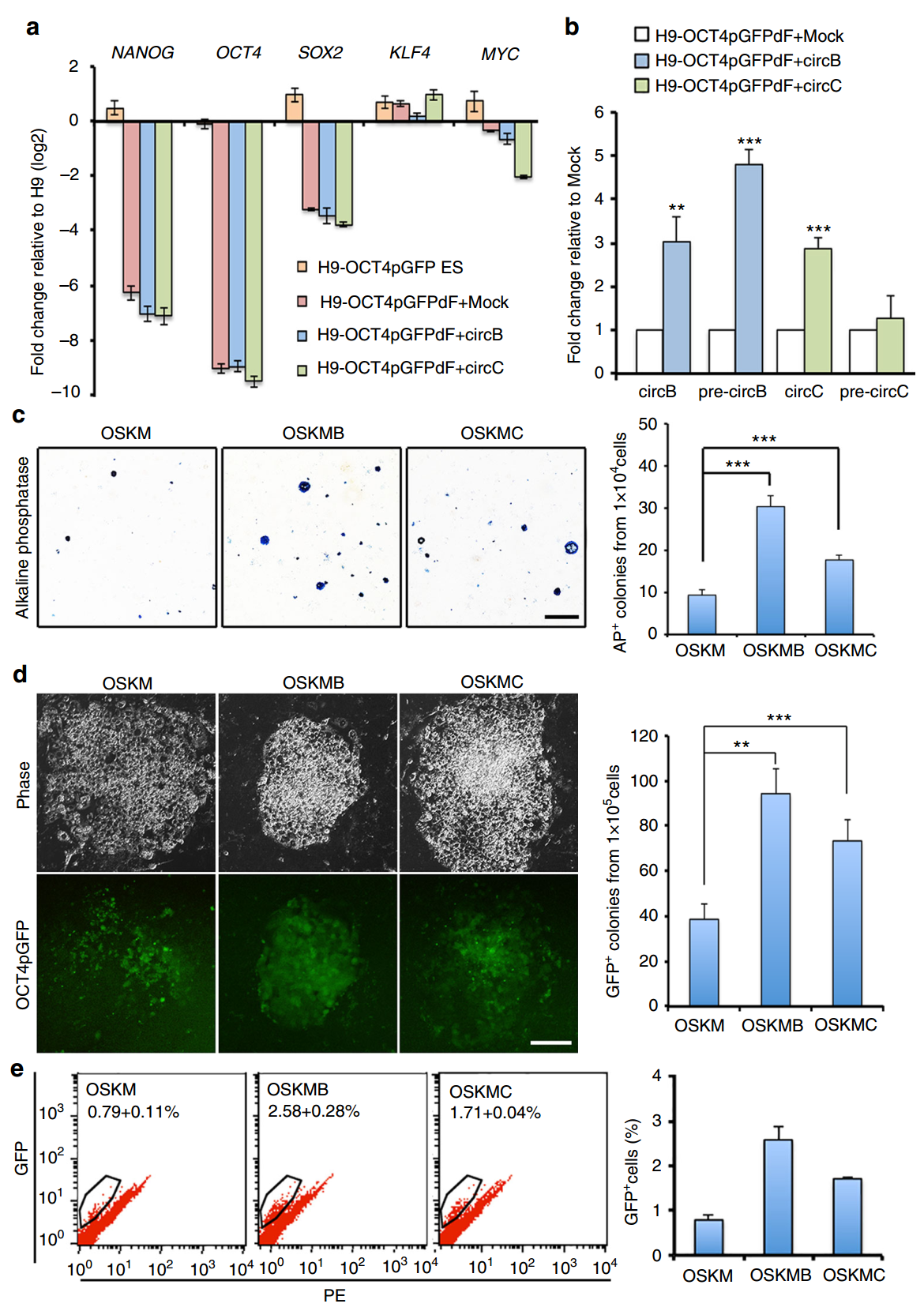
图5 circBIRC6和circCORO1C可促进体细胞重编程效率 (来自[1])
至此,作者基于初步筛选和严格的实验论证,证明了circBIRC6和circCORO1C参与干细胞多能性的调控过程,且过表达它们还可以促进体细胞重编程为iPSC。
circBIRC6和circCORO1C通过竞争性结合miRNA发挥作用
上面的工作详细论证了circBIRC6和circCORO1C在干细胞多能性调控过程中的作用,接下来需要解释它们参与干细胞多能性调控的分子机制。分子机制可以分为两个部分:1.这些目标分子是如何发挥作用的?其下游影响的基因有哪些?2. 这些目标分子是如何产生的?上游调控基因是什么?作者分别针对这两方面进行了探索分析:
circBIRC6和circCORO1C在干细胞中主要影响哪些基因?
竞争性结合miRNA是目前报道最多的circRNA分子机制,作者在本文中也分析了circBIRC6和circCORO1C可能会影响的下游miRNA分子。首先需要分析目标分子的亚细胞定位,结果表明circBIRC6和circCORO1C主要定位于细胞质中。RIP实验表明circBIRC6可结合AGO2而circCORO1C以及其他的线性RNA分子不能结合它们。利用PITA算法预测分析,结合已报到的与细胞多能性的相关性,作者锁定了miR-34a和 miR-145两个miRNA分子。利用3’-生物素标记miR-34a和 miR-145,RNA Pull-down实验证明circBIRC6可以与miR-34a和 miR-145相互作用。荧光素酶报告基因实验也验证了这一作用,并进一步分析了相互作用的序列位点。表达量分析实验表明随着分化进行,circBIRC6逐渐下降,而miR-34a和 miR-145相应升高。
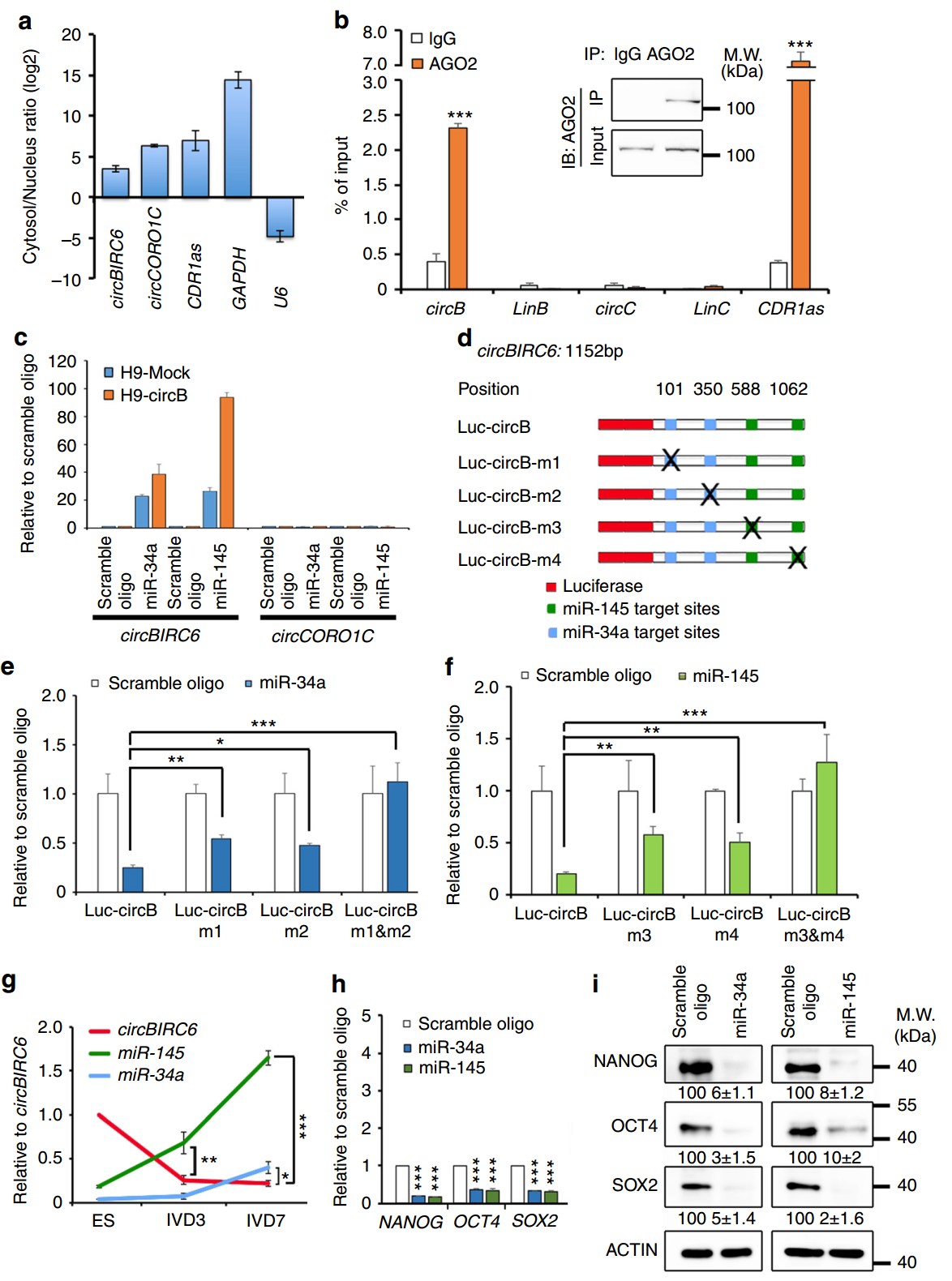
图6 circBIRC6竞争性结合miR-34a和miR-145调控干细胞多能性 (来自[1])
circBIRC6 的形成受到ESRP1调控
RNA剪切因子是调控细胞特异性circRNA形成的重要因素,作者分析了hESC和iPSC中特异性的剪切因子,发现其中ESRP1、SNRPN和HNRNPA1在干细胞中表达,其中ESRP1的表达最高。干扰ESRP1后,circBIRC6表达明显下降,对应的线性的BIRC6表达会升高。ESRP1结合原件呈现GGT-rich的特征,经过分析,在circBIRC6的上游和下游内含子中各发现了一个可能的ESRP1结合位点,还有一个位于circBIRC6的上游区域,作者通过RIP实验证明将预测的circBIRC6上下游的两个结合位点突变之后,ESRP1便无法结合到对应的RNA分子。在干扰ESRP1的细胞中表达circBIRC6的模拟基因,circBIRC6的表达量也明显低于对照组。作者还选择内源基因组中不存在ESRP1结合位点的外显子人为构建两侧插入ESRP1结合位点的报告载体,看ESRP1是否可调控形成circRNA分子,结果表明ESRP1确实可以调控形成circRNA分子。
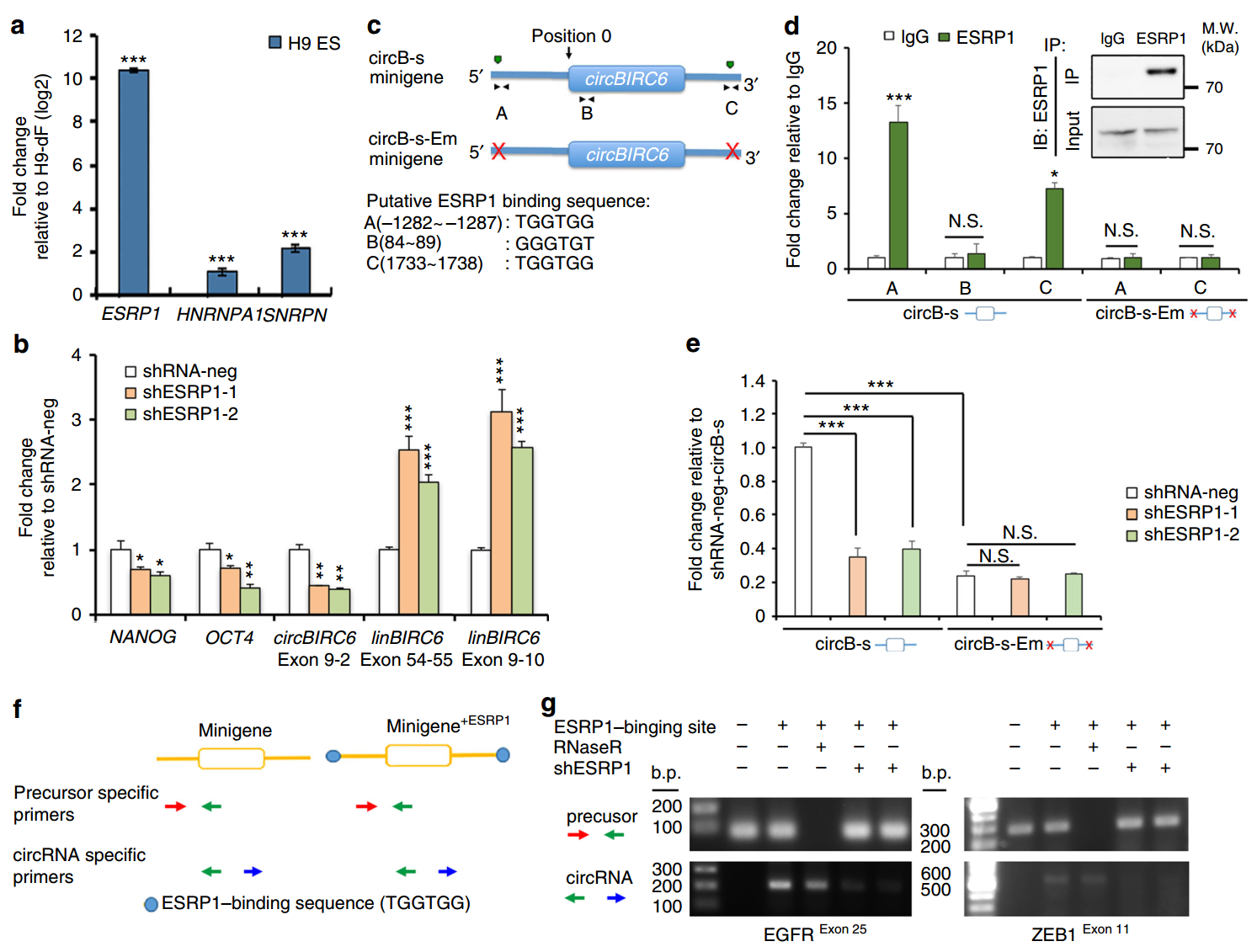
图7 ESRP1调控circBIRC6的形成 (来自[1])
ESRP1在hESC和iPSC中有表达,那么这个基因又是受到什么基因调控的呢?是否与经典的干性相关转录因子有关?作者基于ENCODE ChIP-seq数据分析了ESRP1的启动子区域,发现了该启动子区存在Nanog和Oct4的结合位点,并且存在干细胞特异性的H3K27Ac修饰。通过构建ESRP1启动子区的荧光素酶报告基因,进一步验证了所预测的调控方式。
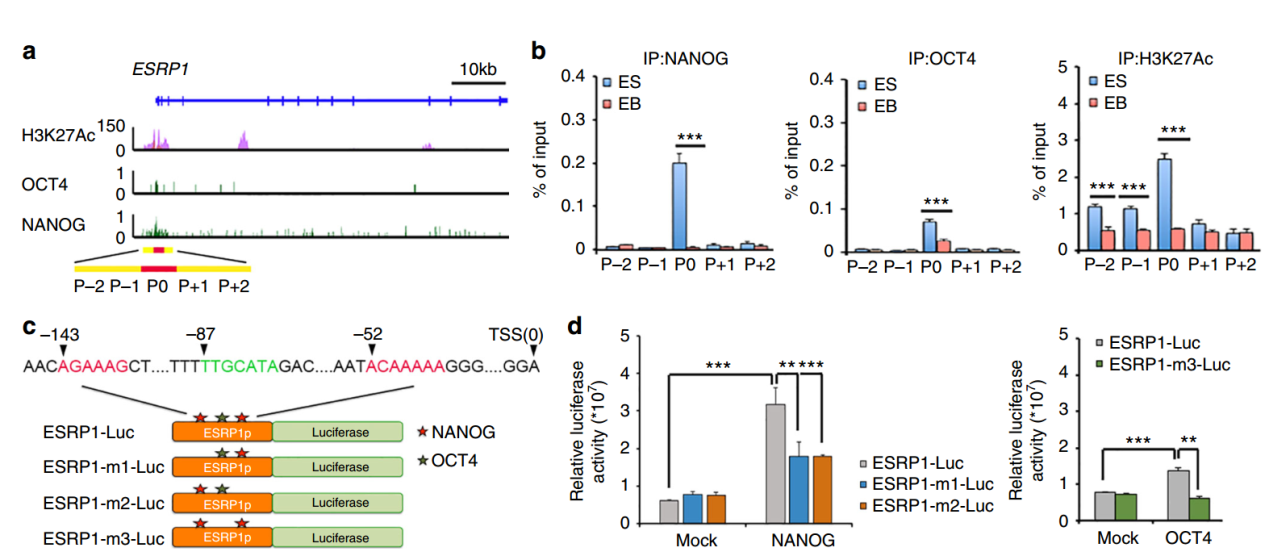
图8 Nanog和Oct4调控ESRP1的表达 (来自[1])
本文首次专门针对干细胞多能性调控相关的circRNA作用机制开展探索,证明了circBIRC6可直接参与多能性的调控,发现和证明其形成过程是受到干细胞特异性的RNA剪切因子ESRP1的调控。本文的相关研究思路和结论拓宽了circRNA功能研究的领域,提高了对circRNA形成机制的理解,尤其是组织/细胞特异性表达的circRNA形成机制的理解,是非常有价值的一项工作,对于组织/细胞特异性circRNA的功能和机制研究均有非常高的参考价值。
参考文献:
1. Chun-Ying Yu, T.-C.L., Yi-Ying Wu, Chan-Hsien Yeh, Wei Chiang, Ching-Yu Chuang & Hung-Chih Kuo, The circular RNA circBIRC6 participates in the molecular circuitry controlling human pluripotency. Nature Communications, 2017.