
德国神经退行性疾病中心的Günter U. Höglinger教授最近在Cell Death& Disease杂志发表题为“Exosomal secretion of α-Synuclein as protective mechanism after upstream blockage of macroautophagy”的研究,病理性α-突触核蛋白聚合物的累积在帕金森疾病中发挥重要的作用。巨自噬是包裹蛋白聚合物进入自噬体后,再融合到溶酶体降解它们的过程。由于作者之前的研究已经发现,药物活化巨自噬保护人神经元细胞免受α-突触核蛋白诱导的细胞毒性,所以猜想抑制巨自噬可能会加重α-突触核蛋白诱导的细胞死亡。但是通过沉默ATG5基因后抑制自噬体的形成,神经元细胞不会遭受α-突触核蛋白诱导的细胞毒性,这可能是在巨自噬被抑制后,存在一种胞内降解或者分泌病理性α-突触核蛋白的补偿途径。沉默ATG5基因并不会影响到泛素蛋白酶系统、分子伴侣系统、分子伴侣街道的自噬和未折叠蛋白反应,却影响到了外泌体分泌α-突触核蛋白的过程。而且阻断了外泌体的分泌会加重α-突触核蛋白诱导的细胞毒性。总之,该研究揭示了在巨自噬被抑制后,神经元细胞会分泌外泌体α-突触核蛋白来减轻胞内的蛋白降解负担,减少了细胞毒性的补偿性保护机制。
帕金森病(PD)是一种以黑质致密部多巴胺能神经元丧失和α-突触核蛋白(α-Syn)在路易小体和Lewy神经突的易受损神经元中积累为特征导致运动障碍的神经退行性疾病。
α-Syn是一种由三个域组成的小突触前蛋白。它的生理功能还没有被研究清楚。编码
α-Syn的基因SNCA发生点突变(如A30P,G51D,A53E,A53T,E46K)会导致常染色体显性的PD。另外,野生型SNCA的二倍体和三倍体也会导致常染色体显性PD。但是,单基因形式的PD是罕见的,而且大多数病例是散发性的。尽管如此,全基因组关联分析一致发现,SNCA的单核苷酸多态性为散发性的PD的主要危险因素。 α-Syn可以由细胞内不同的蛋白质降解机制降解,包括泛素 – 蛋白酶体系统(UPS),伴侣介导的自噬(CMA)或巨自噬(macroautophagy)。
巨自噬,在这里被称为自噬,是一个高度复杂的过程,它始于一个吞噬泡的形成。随后,吞噬泡形成双膜层结构,又称自噬体,是一种在自噬过程中负责吞噬不需要的蛋白质或细胞器的结构。接下来,自噬体融合溶酶体一起形成自噬溶酶体,开始对吞噬的东西进行真正的降解。自噬被认为是一种相当有选择性的过程,具有不同的自噬靶向细胞器(如线粒体自噬)或其他细胞内结构(如脂质吞噬体,聚合物吞噬体)的亚型。之前作者已经证明了药理性刺激自噬可以保护永生化人类多巴胺能神经元前体细胞(LUHMES)免受α-Syn诱导的毒性。于是作者探讨自噬抑制对于α-Syn诱导的细胞死亡的影响。
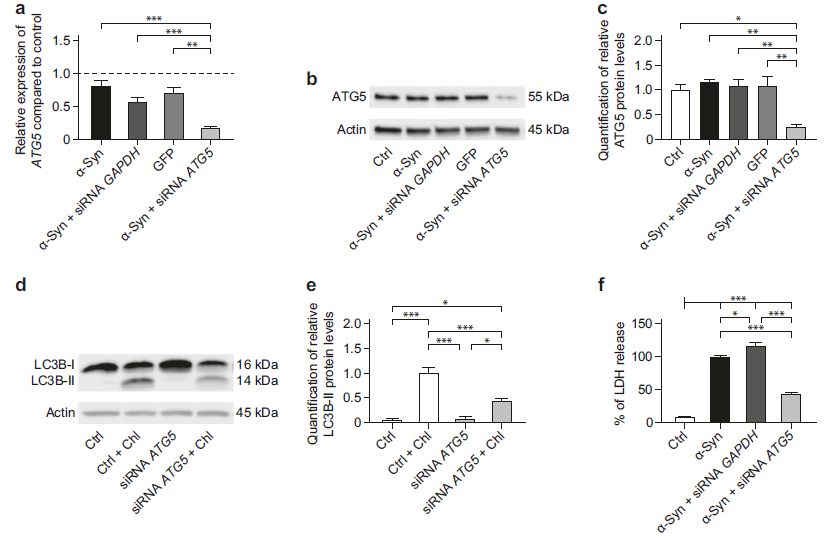
- 首先发现了沉默ATG5可以保护LUHMES神经元细胞免受α-Syn诱导的细胞死亡
相比于(α-Syn过表达组,GAPDH siRNA组,GFP过表达组),(α-Syn过表达+ ATG5 siRNA)组的ATG5的mRNA表达和蛋白水平明显下调(a-c);LC3B-I经磷脂酰乙醇胺连接后变成LC3B-II,作为自噬体的marker,可用于动态观察胞内的自噬流。细胞经氯喹(chloroquine)处理抑制自噬体融合溶酶体后,LC3B-II的蛋白水平明显上调,说明胞内存在正常的自噬流。而ATG5 siRNA+氯喹可以降低LC3B-II的蛋白水平,说明ATG5 siRNA可以抑制自噬体的形成(d-e)。通过定量细胞上清中的LDH释放水平检测ATG5 siRNA对于细胞活性的影响,相比于(α-Syn过表达组,α-Syn过表达+GAPDH siRNA组),α-Syn过表达+ATG5 siRNA组的LDH水平明显降低,代表其细胞毒性明显减小(f)。
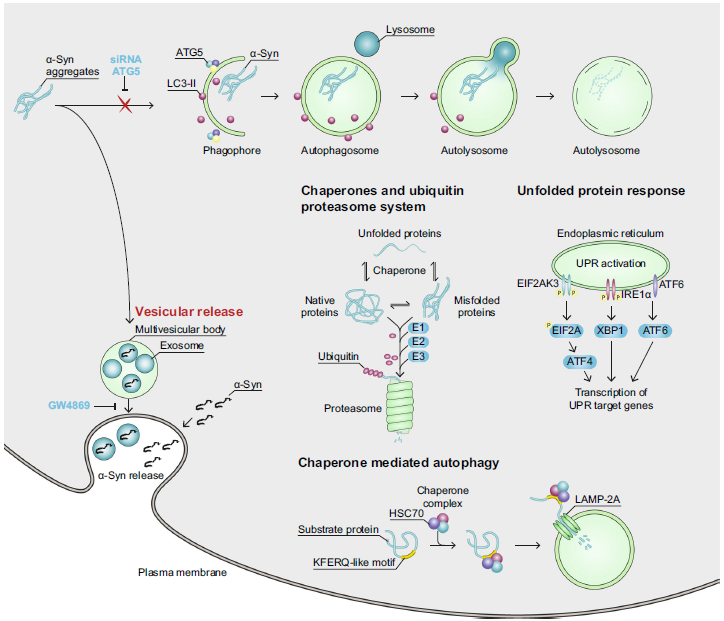
- 作者猜想沉默ATG5基因后,会活化胞内其它的α-Syn处理系统(如UPS、分子伴侣、CMA、UPR和释放α-Syn到细胞上清中)来补偿缺乏自噬降解的机制。
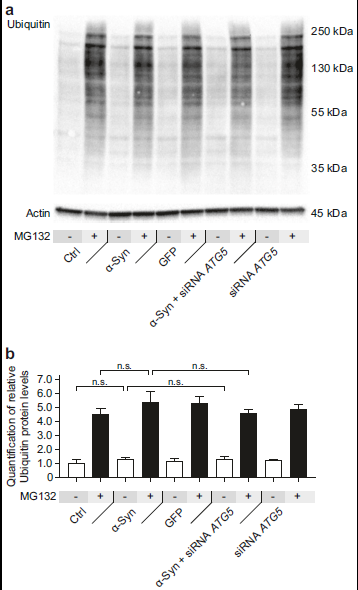
- ATG5基因沉默不会活化泛素蛋白酶体降解UPS途径
给予10 μM MG132处理细胞4h,可抑制UPS,导致泛素蛋白的水平增加。假如α-Syn过表达+ATG5 siRNA组的细胞毒性减小是因为活化了UPS的话,无论给不给予MG132处理,ATG5 siRNA处理都会导致ubiquitin蛋白的水平增加。但是作者发现α-Syn过表达+ATG5 siRNA组的ubiquitin蛋白的水平并没有明显低增加,说明ATG5 siRNA处理并没有活化UPS途径(a-b)。
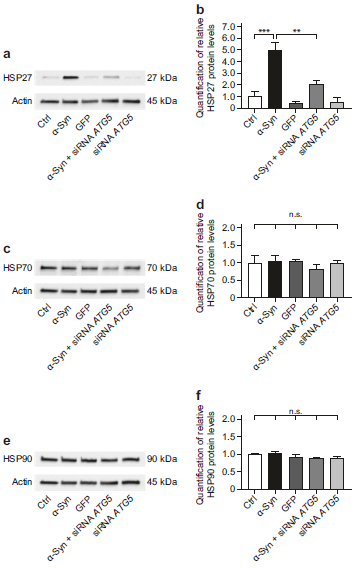
- ATG5基因沉默不会增加补偿性分子伴侣的蛋白水平
分子伴侣靶向不可恢复地错误折叠的蛋白,以通过UPS和自噬途径降解。α-Syn过表达会增加胞内的HSP27蛋白水平,而α-Syn过表达+ATG5 siRNA组的HSP27蛋白水平没有明显的上调(a-b)。相比于其它组别,α-Syn过表达+ATG5 siRNA组的HSP70和HSP90蛋白水平也没有得到明显的上调(c-f)。
- ATG5基因沉默不会增加补偿性CMA相关蛋白水平
含有KFERQ-基序的蛋白质底物可以被细胞溶质伴侣,大小约为70 kDa的热休克同源蛋白HSC70所识别,然后与分子伴侣一起锚定在溶酶体相关膜蛋白质类型2A(LAMP-2A),被内化到溶酶体降解。α-Syn过表达可以增加LAMP-2A 的蛋白水平,而ATG5 siRNA处理并没有在此基础上增加LAMP-2A 的蛋白水平(a-b)。而且各组间的HSC70蛋白水平也没有明显的差异(c-d),进一步说明了α-Syn过表达+ATG5 siRNA组不会活化补偿性CMA相关蛋白。
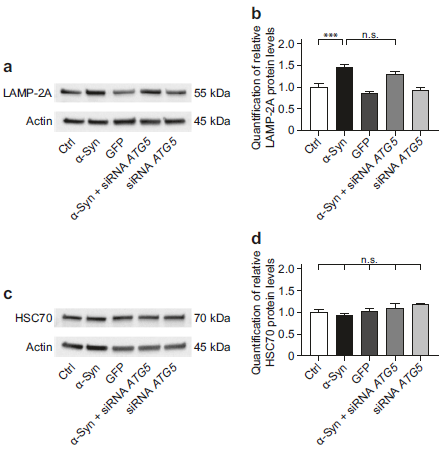
- ATG5基因沉默不会活化补偿性的UPR相关蛋白水平
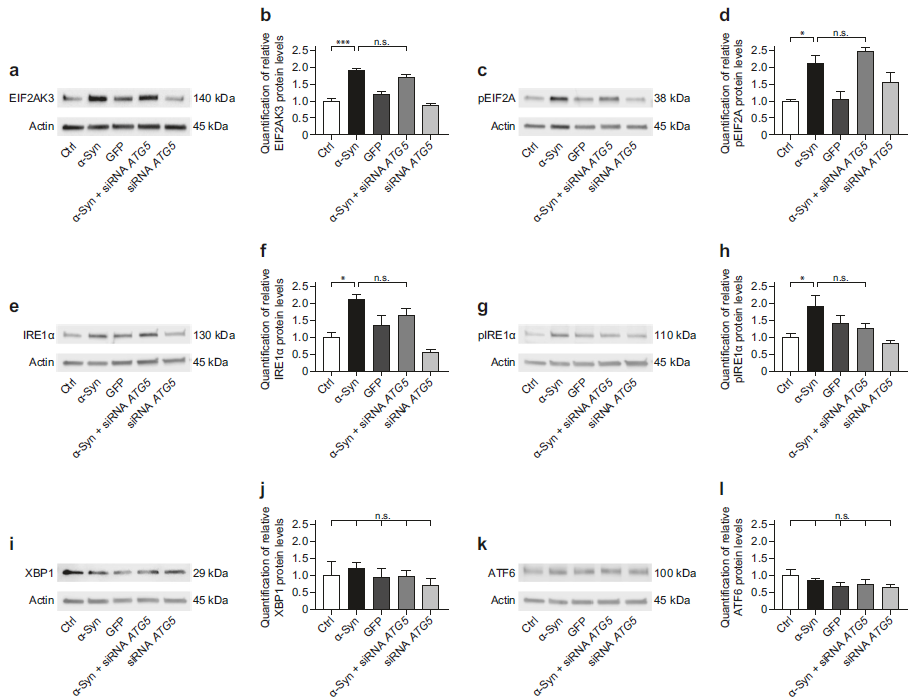
未折叠蛋白反应UPR是一种弃掉错误折叠蛋白的系统,可以被累积在内质网内的错误折叠蛋白所活化,可通过三种应激感应器感应(真核翻译起始因子2 alpha激酶3(EIF2AK3),肌醇必需酶1α(IRE1α)和激活转录因子6(ATF6)。α-Syn过表达可以增加EIF2AK3、pEIF2A 、总IRE1α、pIRE1α的蛋白水平,而ATG5 siRNA处理在此基础上并没有增加EIF2AK3 、pEIF2A、总IRE1α、pIRE1α的蛋白水平(a-h)。α-Syn过表达或者ATG5 siRNA处理都不会影响到XBP1和ATF6蛋白水平(i-l)。这提示α-Syn过表达+ATG5 siRNA处理不会激活补偿性的UPR。
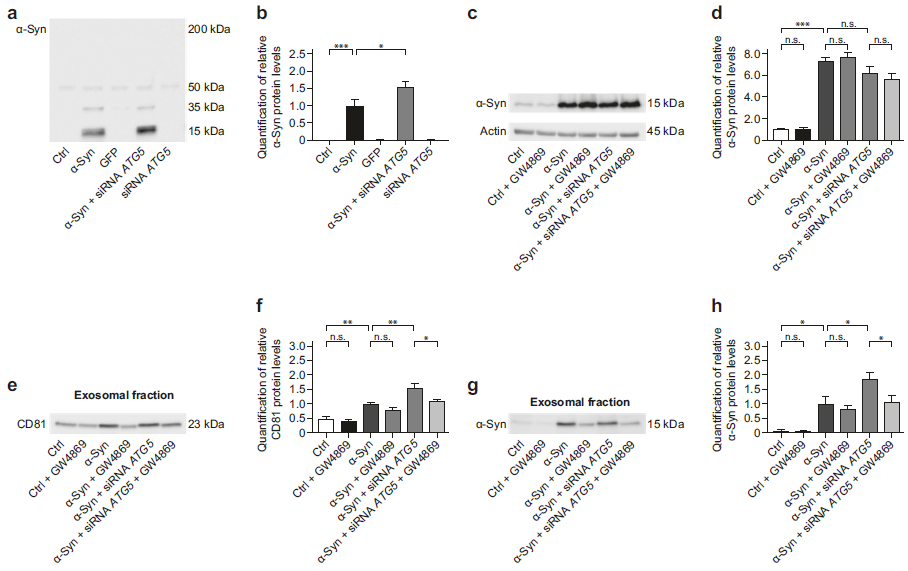
- ATG5 基因沉默会增加外泌体中α-Syn的分泌
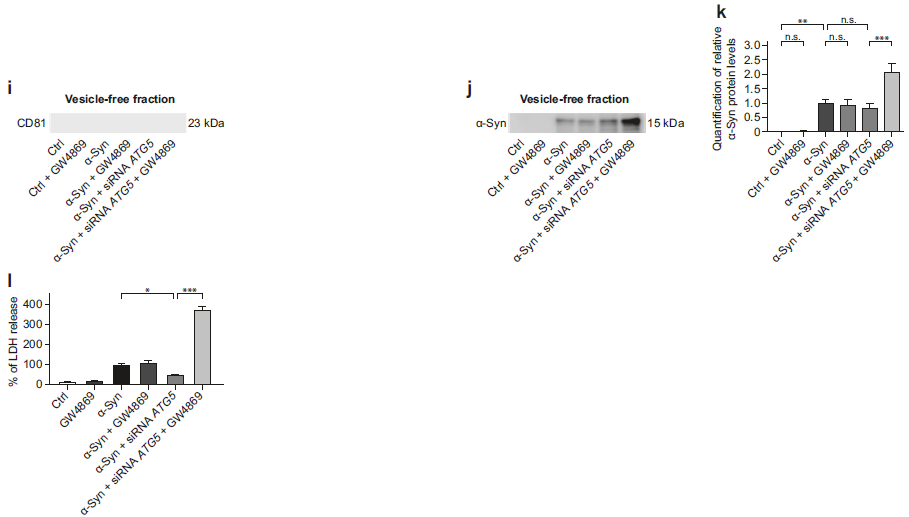
α-Syn过表达四天后可在细胞上清中检测到α-Syn蛋白的分泌,再给予ATG5 siRNA处理后,α-Syn过表达细胞会增加分泌α-Syn蛋白到细胞上清中(a, b);α-Syn过表达会明显增加胞内α-Syn蛋白的表达,但是给予ATG5 siRNA沉默ATG5或者GW4869抑制外泌体分泌后,胞内α-Syn蛋白并没有累积的更多(c, d); α-Syn过表达会明显增加从上清中分离的胞外囊泡部分中的CD81和α-Syn蛋白水平,再给予ATG5 siRNA处理后CD81和α-Syn蛋白水平进一步增加了,提示经过沉默ATG5处理后外泌体的分泌和其中的α-Syn蛋白分泌都增加了,而再同时给予GW4869刺激,这种蛋白进一步增加的现象就消失了(e-h);同时作者也对上清分离的非囊泡部分进行了WB检测,发现无CD81蛋白的存在,说明这部分确实是非囊泡蛋白,而且无论α-Syn过表达处理还是ATG5干扰,都无法增加上清非囊泡部分的α-Syn蛋白分泌,而给予GW4869刺激后α-Syn蛋白分泌增加(i-k),提示抑制自噬后,细胞通过外泌体分泌α-Syn蛋白增加,但当自噬和外泌体分泌都被抑制后,细胞直接释放α-Syn蛋白到上清中。原本沉默ATG5可以减轻α-Syn过表达造成的细胞毒性,但是同时给予GW4869抑制外泌体分泌后,无法减轻α-Syn过表达造成的细胞毒性(l),提示外泌体分泌α-Syn蛋白,对于减轻α-Syn累积造成的细胞毒性是非常的重要。
参考文献
- Fussi N, Höllerhage M, Chakroun T. et al..Exosomal secretion of α-synuclein as protective mechanism after upstream blockage of macroautophagy. Cell Death Dis. 2018 Jul 9;9(7):757. doi: 10.1038/s41419-018-0816-2.