2020年6月19日,德国马克斯·普朗克衰老生物学研究所的Sebastian Gro¨nke和Linda Partridge为共通讯作者在Molecular Cell(IF=15.584)杂志上发表了一篇题为“An Insulin-Sensitive Circular RNA that Regulates Lifespan in Drosophila”的文章,阐述胰岛素信号转导会影响整体circRNA累积,并揭示circSfl在体内衰老过程中的重要作用([1])。

环状RNA(circRNA)在不同物种的神经元中含量丰富,并随着年龄的增长而积累。然而,只有少数circRNA具有功能特征,它们在衰老过程中的作用尚未被揭示。在本篇文章中,作者表征了circRNA与胰岛素介导的寿命延长和衰老之间的功能联系。转录组分析发现,在长寿胰岛素突变果蝇的衰老过程中,circRNA积累减慢。值得注意的是,circSfl在长寿胰岛素突变果蝇中上调,并与胰岛素突变果蝇的寿命延长相关,仅过表达circSfl或其编码的蛋白就足以延长果蝇寿命。
1 在胰岛素突变果蝇的神经元组织中circRNA累积随年龄增长减慢
在各种生物体中,circRNA都会随着年龄的增长而累积,但是circRNA在衰老中的作用还尚不知晓。为了探索circRNA表达随年龄的动态变化,作者对野生型(WDah)果蝇和长寿的三个胰岛素样多肽(dilp 2-3,5)突变果蝇进行全转录水平的深度测序。作者解剖了幼龄(出生后第10天)、中年(出生后第30天)和老年(出生后第50天)果蝇的四个主要成虫组织(脑、有马氏管的肠管、胸部和脂肪体),以组织特异性的方式研究其衰老过程。结果表明,circRNA表达在果蝇大脑中有很明显的倾向,大多数circRNA是大脑所特有的,在其它组织中不表达,这与之前的报道结果相一致。
接下来,作者探索circRNA表达水平随年龄增长的总体变化。如预期的那样,脑组织中总体circRNA水平随着年龄的增长而累积,而在其它组织中,作者仅观察到非常轻度的累积(脂肪体)、无整体变化(肠管)或circRNA水平略微降低(胸部)。令人惊讶的是,circRNA同样在dilp 2-3,5突变果蝇大脑中累积,但比野生型果蝇大脑中的累积程度低得多,表明随着年龄的增长,circRNA的积累会因胰岛素信号的减少而减慢。作者注意到降低的胰岛素信号转导也会影响其它组织中各个时间点的总体circRNA水平,但是它们并没有一致的变化,如在肠管组织中circRNA水平升高,在胸部组织中circRNA水平降低。总之,作者观察到,不仅在组织特异性方面而且在时间特异性方面,circRNA的表达同样存在显着差异,这表明circRNA在衰老过程中具有重要作用。
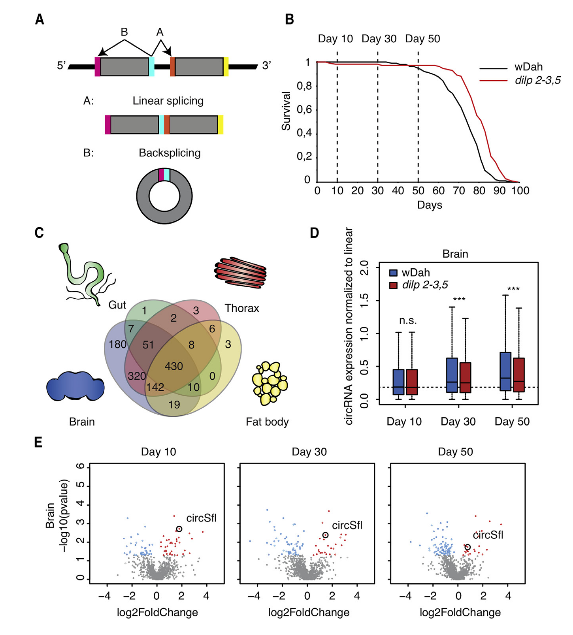
图1 衰老过程中长寿胰岛素突变果蝇的组织特异性circRNA表达谱
2 circSfl在不同的长寿胰岛素突变果蝇中上调
接下来,作者寻找独立于宿主基因并特异性地调控dilp 2-3、5突变果蝇的circRNA,因为,这可以表明circRNA的调控是特异性的,而不仅仅是与宿主基因共同发挥作用。大部分差异表达的circRNA是在dilp 2-3、5突变果蝇的大脑和脂肪体内检测到,仅有少数存在于肠管和胸部组织中。作者确定由sulfateless(sfl)基因编码的circRNA circSfl是果蝇体内表征的主要靶标,因为circSfl在果蝇的几乎所有组织和所有时间点都表达并在胰岛素突变果蝇中上调,此外,它还是最强的调控性circRNA之一。接下来,作者进行RNase R处理证实circSfl对RNase R具有抗性。随后,Sanger测序验证了circRNA特异性反向剪接区域的存在,表明circSfl确实是一个环状转录本。
为了验证RNA测序结果,作者使用特异性引物通过qRT-PCR来分析circSfl和源于Sfl基因的两个不同线性转录本(RA和RB)的表达水平,RA和RB的不同之处在于RA基因的5’非翻译区(UTR)有一个37bp长的功能未知的外显子。结果表明,在dilp 2-3、5突变果蝇的所有组织和时间点中,circSfl的表达均被上调。有趣的是,Sfl RB的表达,而不是RA,与dilp 2-3、5突变果蝇中的circSfl表达模式相关,暗示线性和环状同系物的剪接之间可能存在联系。与之一致的是,circSfl在其它长寿胰岛素突变果蝇中也同样上调:遗传性切除正中神经分泌细胞(MNC),MNC可以产生胰岛素;在肌肉中过表达dFoxo。在这两种情况下,circSfl的表达均被上调,而非线性转录本Sfl RA和RB。有趣的是,MNC切除果蝇中circSfl的上调也取决于dFoxo,它是MNC切除果蝇延长寿命所需的胰岛素信号传递的下游转录因子。在野生型和MNC切除果蝇中,敲低dFoxo均会下调circSfl的表达。值得注意的是,在饲喂可延长寿命的雷帕霉素果蝇中circSfl表达并没有增加,在饮食限制下的果蝇甚至在mth1突变果蝇中circSfl表达也仅仅是轻微下调。这些结果表明,circSfl在长寿胰岛素突变果蝇中特异性地上调,而这种上调取决于转录因子dFoxo,它是减少胰岛素信号传递以延长寿命所必需的。
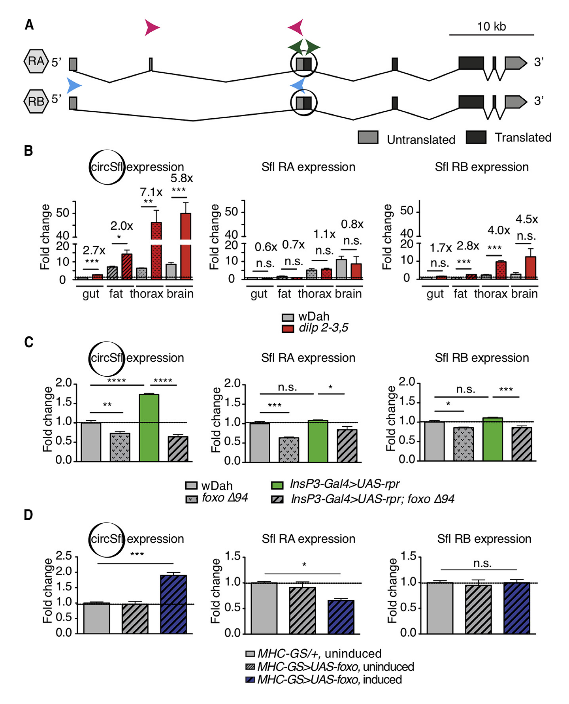
图2 circSfl以dFoxo依赖性的方式在长寿胰岛素突变果蝇中上调
3 构建circRNA过表达突变体
下一步,作者的目标是构建circSfl的过表达突变体。作者测试三种不同构建体的circSfl过表达能力:circSfl-exon(仅含有产生circSfl的外显子)、circSfl-1,000(circSfl外显子加上两侧1,000 bp的内含子)和circSfl-inverted(circSfl-inv;合成circSfl外显子上游1000 bp内含子的反向互补序列,并人工的添加到circSfl外显子下游),仅仅只有circSfl-inv构建体可以显著过表达circSfl,而Sfl RA和RB的表达水平并没有受到很大影响,以下将circSfl-inv称为circSfl过表达突变体。为了验证这种过表达系统是否也适用于其它circRNA,作者为circBtsz合成了类似的过表达构建体。与circSfl的结果一致,circBtsz-inv突变体同样强烈过表达circBtsz,而circBtsz-exon和circBtsz-1000均不影响circBtsz的表达水平。总之,作者利用工程性反向互补的侧翼内含子,来在果蝇体内强烈过表达特异性的circRNA。
4 过表达circSfl可以延长果蝇寿命
由于circSfl在不同的长寿胰岛素突变果蝇中持续上调,作者想知道仅仅过表达circSfl是否足以延长果蝇寿命。在低胰岛素条件下,circSfl在果蝇的四个测试组织中均上调,其中,在肌肉和脑组织的上调最显著。因此,作者选择神经元特异性的elav-Gal4和肌肉特异性的MHC-Gal4来驱动特定组织过表达circSfl,并选择da-Gal4来普遍过表达circSfl。引人注目的是,在这三个Gal4驱动程序中过表达circSfl均会导致雌性果蝇寿命显著延长。在肌肉组织、神经元和普遍过表达circSfl中,果蝇的平均寿命分别提高了12%、4.5%和15%。其中,普遍过表达circSfl的果蝇的寿命延长最为显著。相比之下,雌性果蝇体内NP1-Gal4驱动肠道特异性过表达circSfl或雄性果蝇中普遍过表达circSfl并没有显着改变果蝇的寿命,表明circSfl对寿命的延长存在组织特异性和性别特异性。值得注意的是,qRT-PCR表明,在上游激活序列(UAS)对照果蝇(UAS-circSfl /+)中circSfl的表达较低,而这些果蝇与Gal4对照果蝇相比寿命并不长,这表明在野生型背景下,circSfl的强表达是延长寿命的必要条件。有趣的是,尽管过表达circSfl会延长果蝇寿命,但与胰岛素突变果蝇不同,它们并未显示出抗压或爬升能力的增强、果蝇较小以及发育迟缓。这提示circSfl调节一种机制,该机制专门影响寿命,但并不影响胰岛素信号减少的其它多效性后果。综上所述,作者的研究表明,circRNA在衰老过程中发挥重要作用,它可以延长寿命,这有助于了解circRNA在体内的功能。
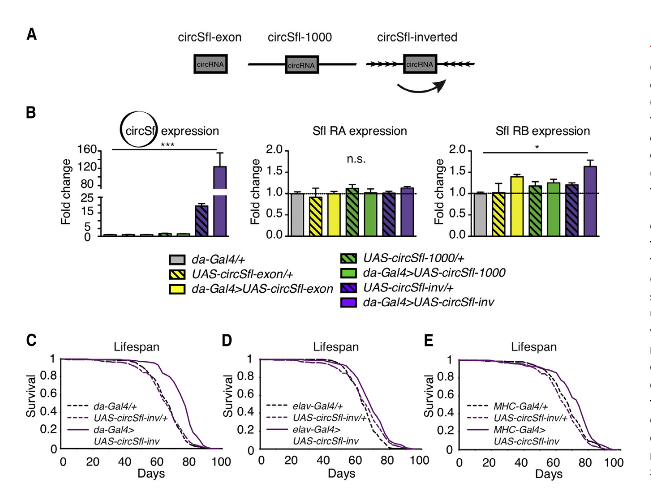
图3 过表达circSf1可延长寿命
5 胰岛素介导的寿命延长依赖于Sfl-RA和circSfl
由于过表达circSfl足以延长果蝇寿命,作者想知道circSfl是否也是延长胰岛素突变果蝇寿命所必需的。因此,作者设计并合成了靶向circSfl反向剪接区域的siRNA来敲低circSfl。然而,所测试的三个siRNA均不能显著降低circSfl的表达,也不能影响线性转录本RA和RB的水平。这可能是因为,siRNA设计非常局限于circRNA所特有的反向剪接区域,导致设计的siRNA并不是总能够成功地敲低。值得注意的是,作者发现在长寿dilp 2-3,5突变果蝇中,circSfl的反向剪接与线性Sfl RB转录本(而非RA)的选择性剪接相关。随后,作者想要通过影响宿主基因的选择性剪接来改变circSfl的表达。为了阐明circSfl反向剪接和线性Sfl RB转录本选择性剪接之间的联系,以及是否可以通过改变选择性剪接来影响circSfl表达,作者通过CRISPR-Cas9生成了Sfl∆ex2突变果蝇,Sfl∆ex2突变果蝇在Sfl 5’UTR上缺失约250 bp,包括37 bp长的外显子2,这是Sfl RA所特有的。因此,Sfl∆ex2突变果蝇仅表达Sfl RB。qRT-PCR表明,在Sfl∆ex2和dilp 2-3,5突变果蝇中,总的线性Sfl转录本水平并没有受到影响,正如预期的那样,在Sfl∆ex2突变果蝇中检测不到Sfl RA,但是Sfl RB的表达却增高了4倍以上,额外地弥补了Sfl RA的缺失。此外,在Sfl∆ex2突变果蝇中circSfl的表达降低了50%,而在dilp 2-3,5突变果蝇的基础上,Sfl∆ex2突变会消除circSfl表达的上调。这些结果表明,circSfl 在dilp 2-3,5突变果蝇中的上调取决于Sfl RA,并证明circSfl的反向剪接和线性Sfl转录本的选择性剪接之间确实存在联系。
Sfl∆ex2突变体可用作circSfl敲低,因为在这些突变体中circSfl的表达有所降低。因此,作者表征了Sfl∆ex2突变果蝇以及Sfl∆ex2和dilp 2-3,5双突变果蝇,以测试Sfl∆ex2和dilp 2-3,5之间的遗传上位性。有趣的是,虽然Sfl∆ex2单突变果蝇显示出正常的产卵行为,但在Sfl∆ex2和dilp 2-3,5双突变果蝇中,dilp 2-3,5介导的产卵减少得以部分恢复。同样,较小的dilp 2-3,5突变果蝇部分被Sfl∆ex2和dilp 2-3,5双突变恢复,这表明dilp 2-3,5突变果蝇的繁殖力和大小表型部分取决于Sfl RA和circSfl。相反,Sfl∆ex2突变果蝇发育延迟,与dilp 2-3,5突变果蝇相比,Sfl∆ex2和dilp 2-3,5双突变果蝇的发育时间更长,表明发育延缓是受独立的机制调控。最后,Sfl∆ex2和dilp 2-3,5双突变果蝇的寿命显著降低,相比于dilp 2-3,5突变果蝇。总之,作者证明了果蝇的减小大小、降低功能性和延长寿命,而不是延迟dilp 2-3,5突变果蝇发育,取决于Sfl RA和circSfl。
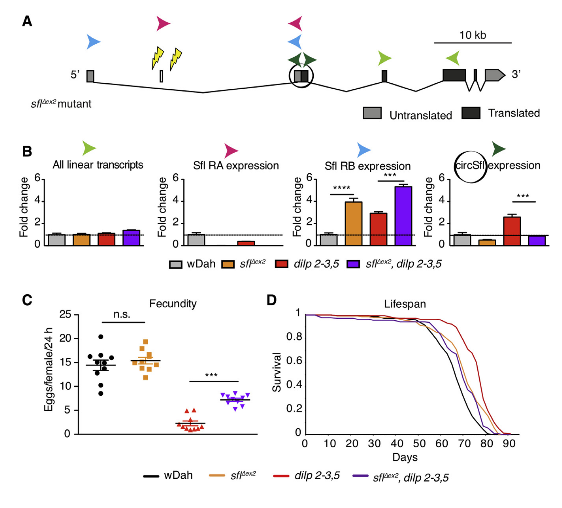
图4 circSfl的反向剪接取决于上游非编码外显子的存在,这对于胰岛素突变介导的寿命延长至关重要
6 circSfl可以被翻译和动态调节
作者已经证明了circSfl在体内的功能,过表达circSfl足以延长果蝇寿命,并且circSfl的产生与宿主基因的选择性剪接相关。然而,circSfl的分子功能仍然未知。一些circRNA被认为可以海绵吸附microRNA(miRNA),例如,人类CDR1as含有大约70个miR-7的结合位点。相反,在circSfl序列中,作者只鉴定出可以与9种不同miRNA相结合的10个miRNA结合位点,这与果蝇的其它circRNA相比,结合位点相对较少,因此circSfl不太可能充当miRNA海绵。最近有研究表明,当某些circRNA携带其宿主基因的起始密码子并在反向剪接连接区域后存在终止密码子时,它们就可以被翻译成蛋白质。而circSfl正好满足这两个条件,表明它具有蛋白编码潜力。如果circSfl可以编码蛋白质,那么它与传统线性转录本编码的Sfl全长蛋白有相同的N端,包括完整的跨膜结构域,但缺少Sfl全长蛋白C端的酶活性结构域。为了确定circSfl RNA转录是否与其翻译机制相关,作者分析了circSfl在多核糖体谱中的存在情况,并与野生型和dilp 2-3,5突变型脂肪体的总RNA测序结果进行比较。作者观察到,包括宿主基因ATG起始密码子的circRNA在多核糖体中富集;例如,circCamKI、circPkn和circPdk1。此外,在野生型和dilp 2-3,5突变型脂肪体中,circSfl是多核糖体最富集的circRNA之一。
为了测试circSfl是否真的在体内编码蛋白,作者使用CRISPR-Cas9生成FLAG::Sfl突变果蝇。将FLAG标签插入内源性Sfl基因,在N末端标记宿主基因生成的全长Sfl蛋白,以及可能由circSfl产生的蛋白。使用western blot分析FLAG :: Sfl突变果蝇,作者观察到两个不同的条带,分别对应于分子量约110 kDa(Sfl全长)和25 kDa。根据序列,circSfl ORF(包括FLAG标签)翻译会产生24.75-kDa的蛋白。但是,25kDa的条带也可能来自未知的较短的Sfl异构体或Sfl蛋白降解产物。为了排除这两种可能,作者构建一个仅过表达FLAG标记的circSfl,且不改变FLAG标记的全长Sfl蛋白水平的突变体(称为FLAG :: circSfl)。Western blotting实验表明,过表达FLAG :: circSfl会导致25 kDa的条带加深。此外,作者注意到,与FLAG :: Sfl突变果蝇相比,该蛋白带在SDS-PAGE上的移动速度更快,表明该蛋白的尺寸较小,或是来自FLAG :: Sfl突变果蝇的蛋白具有不同的翻译后修饰。为了进一步证实该蛋白条带确实对应于circSfl编码的蛋白,而不是传统的蛋白降解产物。作者使用UAS-Sfl果蝇,具有N端血凝素(HA)标签并可以表达Sfl的cDNA,它可以生成线性Sfl转录本,而非circSfl。蛋白质印迹分析显示,只有一个蛋白条带与Sfl全长蛋白相对应,即使在elav-Gal驱动的过表达Sfl果蝇中,也没有观察到25 kDa的条带。
作者试图推测circSfl蛋白可能在胰岛素介导的寿命延长中起作用。蛋白印记实验显示,circSfl蛋白水平确实在dilp 2-3,5突变果蝇中上调。最后,为了确定circSfl编码的蛋白是否足以延长果蝇寿命,作者构建了一个从线性转录本中过表达circSfl ORF的转基因果蝇系。值得注意的是,线性circSfl ORF的普遍表达显着延长了雌蝇寿命。因此,多条证据表明circSfl可以编码蛋白,并且该蛋白足以延长果蝇寿命。
图5 circSfl编码的蛋白在胰岛素突变果蝇中上调
7 Sfl的神经元特异性过表达可以延长果蝇寿命
如果circSfl编码小蛋白,则该蛋白与传统线性转录本编码的Sfl全长蛋白有相同的N端。Sfl线性转录物可以编码一个N-脱乙酰基酶/ N-磺基转移酶(Ndst),该酶通过高尔基体中N-乙酰氨基葡糖(GlcNAc)的N和6-O位置的硫酸化催化硫酸乙酰肝素(一种糖胺聚糖)的合成。有趣的是,circSfl ORF覆盖了Sfl的完整胞质N末端以及疏水性跨膜结构域,但缺乏对硫酸乙酰肝素合成重要的光酶结构域。由于过表达circSfl蛋白可以延长果蝇寿命,所以作者接下来测试过表达Sfl全长蛋白是否也影响果蝇寿命。作者分别在UAS-Sfl转基因果蝇的肌肉、肠道和神经元中过表达Sfl,发现在肌肉或肠道中过表达Sfl不会影响果蝇寿命,而普遍过表达Sfl对于果蝇的寿命是有害的。然而,使用elav-Gal4驱动雌性果蝇神经元特异性地过表达Sfl,会使其寿命延长约15%。与过表达circSfl的结果相一致,过表达Sfl并没有增加雄性果蝇的寿命。令人惊讶的是,在dilp 2-3、5突变果蝇中,神经元特异性过表达Sfl会降低果蝇寿命,指出Sfl和dilp 2-3,5之间在遗传上存在相互作用。类似于circSfl过表达突变体,作者在过表达Sfl的果蝇中并没有观察到饥饿压力抗性、攀爬能力、体重或发育的任何差异。相比之下,作者观察到da-Gal4驱动普遍敲低Sfl在幼虫后期的发育是致命,而在成虫阶段会导致寿命缩短。这些结果表明,circSfl和Sfl既有相似功能,又有独立作用,这与circSfl蛋白存在Sfl全长蛋白相同的完整胞质N末端和跨膜结构域,在高尔基体中缺乏C末端的观察结果相一致。
图6 神经元特异性过表达Sfl可以延长果蝇寿命
8 Dally作为Sfl延长寿命的下游靶标
Sfl可以介导硫酸乙酰肝素的合成,果蝇中存在四种Sfl靶向的硫酸乙酰肝素蛋白聚糖:Dally、Dally-like(Dlp)、Syndecan(Sdc)和Perlecan。为了验证Sfl是否通过硫酸乙酰肝素蛋白聚糖来延长寿命,作者在神经元中过表达Dally、Dlp和Sdc。令人惊讶的是,只有过表达Dally对寿命的影响与过表达Sfl对寿命的延长趋势相一致,表明Dally可能是过表达Sfl延长寿命的下游靶标。为了进一步验证,作者进行遗传上位实验,并使用RNAi技术来敲低过表达Sfl果蝇中的Dally。结果表明,敲低Dally废除了Sfl诱导的寿命延长,这表明Dally是Sfl调节寿命的重要的下游靶标。总之,本研究表明在胰岛素突变果蝇中circSfl和Sfl的线性剪接被改变,并且过表达circSfl或Sfl可以以组织特异性的方式延长寿命。
图7 硫酸乙酰肝素蛋白聚糖Dally介导Sfl诱导的寿命延长
参考文献[1]:Weigelt CM, Sehgal R, Tain LS, et al. An Insulin-Sensitive Circular RNA that Regulates Lifespan in Drosophila [published online ahead of print, 2020 Jun 19]. Mol Cell. 2020;S1097-2765(20)30396-8. doi:10.1016/j.molcel.2020.06.011