引 言
合成信使RNA已在基于mRNA的治疗及疫苗开发过程中成为了一种新兴的应用技术。近年来研究发现,环状RNA(circRNAs)能在细胞中具备良好的稳定性和持久的蛋白表达能力,合成环状RNA作为一种新型的mRNA合成技术显现出来优越的应用潜能。然而,环状RNA的翻译控制仍具有挑战性。
2023年1月,Nucleic Acids Research杂志上发表了一篇文章Synthetic circular RNA switches and circuits that control protein expression in mammalian cells。作者开发了“circRNA开关”,能够通过感知细胞内RNA或蛋白质来控制circRNA的蛋白表达。其方法在于:将miRNA结合或蛋白结合序列分别插入非翻译区(utr)或柯萨奇病毒B3内部核糖体进入位点(CVB3 IRES),设计成了miRNA及蛋白响应的环状RNA翻译开关,工程化的circRNA在不诱导严重细胞毒性和免疫原性的情况下有效表达蛋白,并对靶mirna或蛋白做出反应,从而控制circRNA的翻译水平。此外,作者还构建了“基于circRNA的基因电路”,通过检测内源性miRNA及蛋白响应的circRNA,从而选择性地激活翻译。本项研究为工程化RNA的设计提供了新的见解,并在RNA合成生物学和基因细胞治疗领域具有广泛的潜力。

01
circRNA的环化设计及测试
作者选择经过人工改造的内含子-外显子(PIE)自剪切环化系统。结果显示,通过PIE系统可体外人工制备circRNA(图1),制备的环状RNA在不引起严重细胞毒性和免疫原性的情况下可高效表达靶蛋白(图2)。
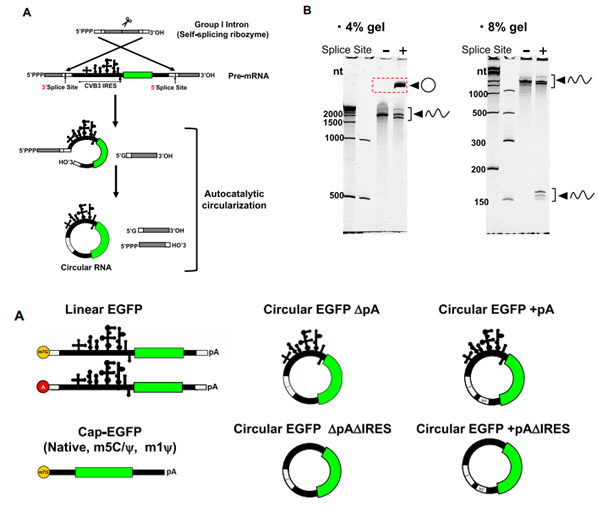
图1. circRNA制备设计
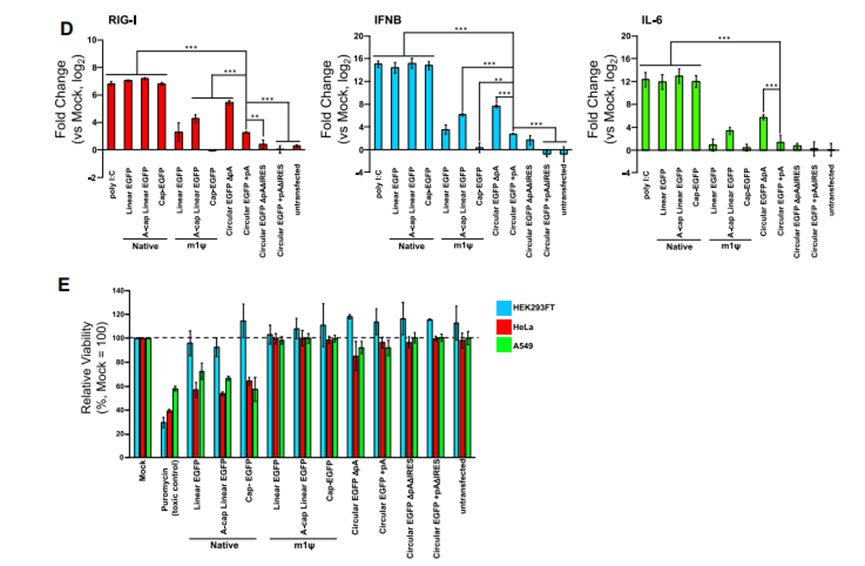
图2. circRNA免疫原性及翻译能力测试
02
miRNA响应的circRNA开关构建及测试
作者设计了circRNA(在utr处含有完全互补的anti-miR序列),并通过共转染各种miRNA模拟物对其进行评估。通过在CVB3 IRES序列(5,端)之前或egfp编码序列(3,端)之后插入anti-miR,制备了4种不同的人类miRNA(hsa-miR-206,hsa-miR-302a-5p,hsa-miR-21-5p和hsa-miR339-5p)响应的circRNA开关。转染circRNA开关和相应的miRNA模拟物24h后,通过流式细胞仪和荧光显微镜分析circRNA的EGFP表达。结果发现,所有设计的miRNA响应性开关均通过感知靶miRNA模拟物来抑制EGFP表达(图3A)。观察到的ON状态和OFF状态之间的倍数变化(约2 ~ 38倍范围)取决于靶miRNA、anti-miR插入位置和内部polyA序列的存在与否(图3B)。作者推测miR21-5p和miR-339-5p响应开关的相对低倍数变化可能是由于HEK293FT细胞中内源性miRNA活性介导的。因此,作者在细胞中添加靶向miRNA抑制剂来阻断内源性miRNA的活性。结果发现,共转染miR-21-5p抑制剂可挽救circRNA翻译;而miR-339-5p抑制剂未观察到这一情况。这一结果表明,在miR-21-5p响应开关的情况下,观察到的低倍数变化是由于内源性miR-21-5p活性弱化了ON状态。此外,作者还研究了circRNA构建体的开关是否可以检测内源性miRNA并调节靶细胞的翻译,重点关注HEK293FT中的hsa-miR-17-5p和HeLa和A549中的hsa-miR-21-5p,因为这些miRNA在每种细胞类型中都有效表达。结果表明,miRNA响应的circRNA开关能有效地检测靶miRNA,并控制其翻译水平。
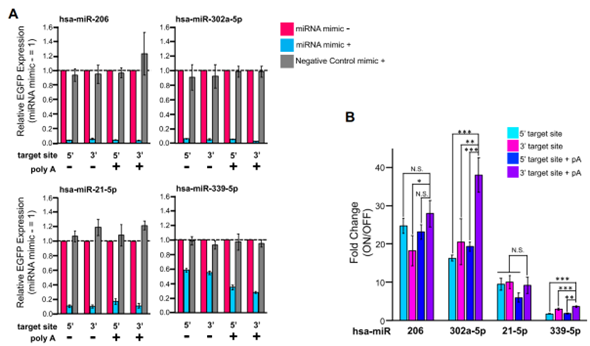
图3. miRNA响应的circRNA开关的构建及测试
03
改造CVB3 IRES元件使其
成为RBP响应的circRNA开关
作者利用两个RBP:MS2噬菌体外壳蛋白(MS2CP)和剪接体相关SNRPA(U1A)蛋白设计了蛋白质响应的circRNA。为了研究蛋白质结合的部位能够使IRES依赖的翻译受到抑制,作者测试了4个CVB3 IRES变异体(变异体1-4),这些变异体是通过插入蛋白结合基序(MS2SL和U1A适配子)设计的,同时参考了翻译起始所需的二级结构模型(如图4A)。插入MS2CP或U1A结合motif表明,只有将motif插入到结构域VI (MS2CP)或VII (U1A)中的变异体4中,才能够在靶RBP存在的情况下抑制翻译,而其他变异体不能有效表达报告基因或对靶RBP无应答(如图3B)。根据这些结果,可以预测翻译起始因子eIF4G和eIF4A(与结构域V-vii结合)的结合位点附近或下游的CVB3 IRES插入的motif,或针对18S rRNA的反义区域(结构域V和VI之间的连接区域),这是可能有效产生RBP响应的circRNA开关(如图4C)。为了优化插入位置,进一步设计了变异体5和6,其插入motif位于结构域VII茎环下游的20 nt或98 nt(仅位于ORF上游),并比较了它们与变异体4的抑制效率。结果表明,存在MS2CP或U1A的情况下,MS2SL的变异体4和U1A的变异体5的翻译抑制效果最好。作者应用这些变异体来评估它们在circRNA中的表现,以确认内部polyA序列的存在或缺失对翻译抑制和倍数变化的影响。如在存在或不存在MS2CP或U1A的情况下,在circRNA开关结构中观察到的翻译抑制均有效。总之,这些结果表明,RBP响应的circRNA开关可以通过CVB3 IRES的结构域VI和VII区域来设计。
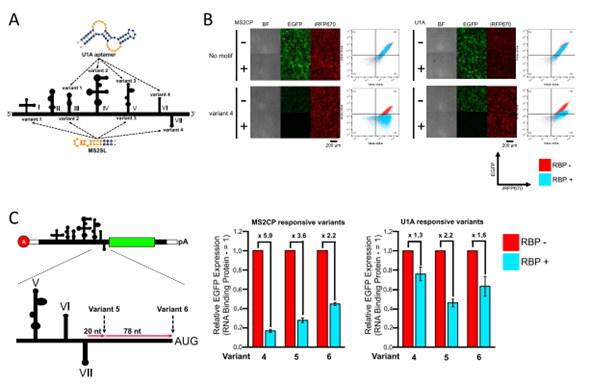
图4. CVB3 IRES工程化改造为RBP响应的circRNA开关
初稿:青栀
审核:刘弋
指导:山人
原文链接:
https://doi.org/10.1093/nar/gkac1252
转载请联系邮箱授权:circRNA@163.com