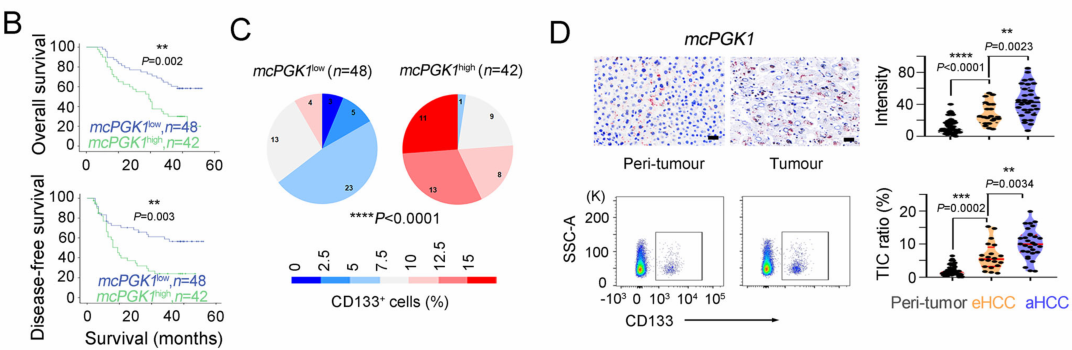
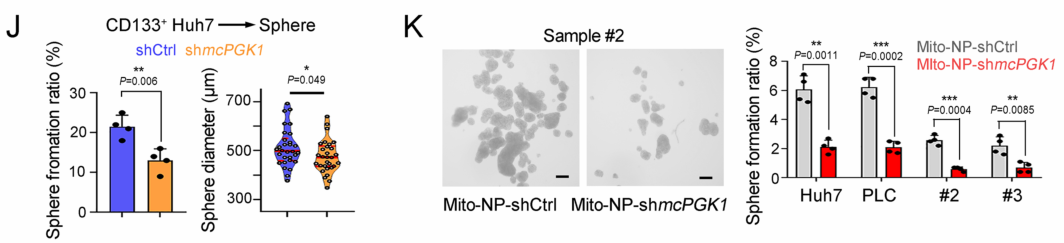
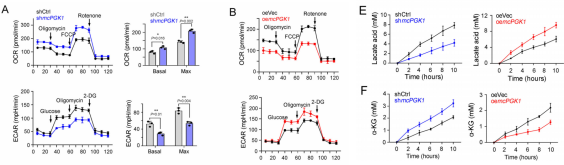
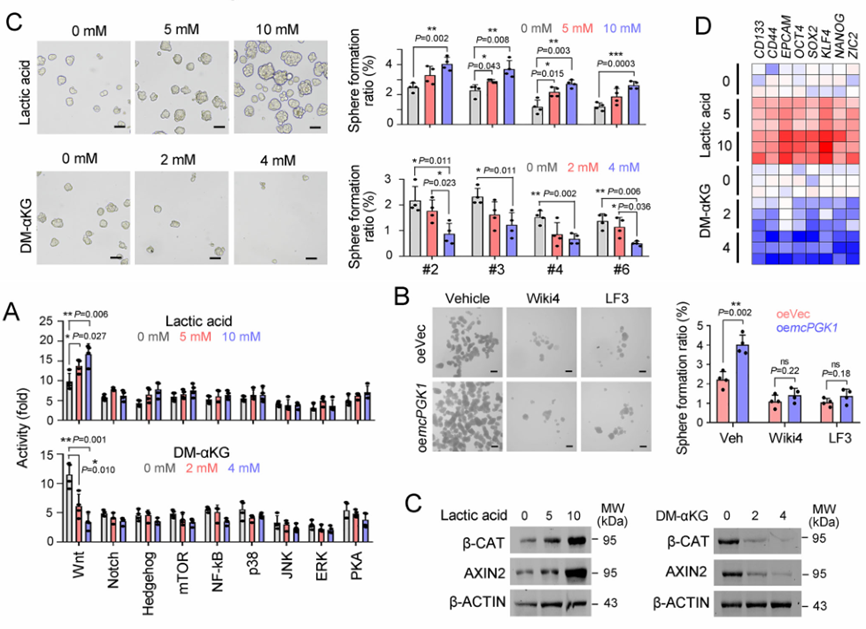
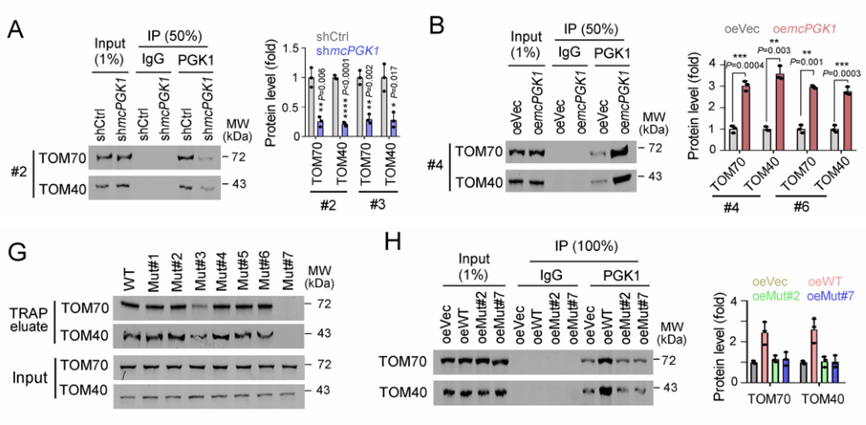
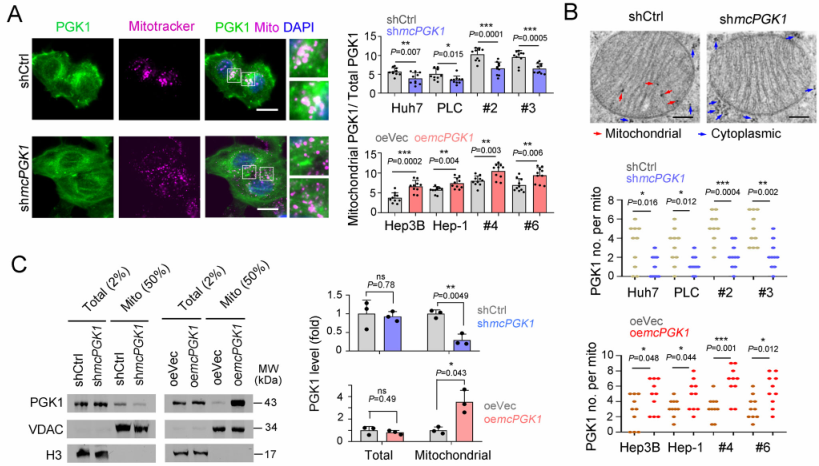
肝癌是一种常见的肿瘤类型,由于其异质性,导致很多肝癌患者预后效果极差。肝脏肿瘤起始细胞(TICs)有助于肿瘤的转移、生长和耐药。代谢改变是癌症的标志,在肝脏肿瘤发生过程中起着重要作用。然而,代谢改变在TIC中的作用仍未得到充分的研究。环状RNA (circRNAs)是一种新发现的调控RNA分子,在多种生物过程中发挥着重要的调节作用,如作为miRNA的分子海绵、调节因子和发挥RBP功能等等[1]。越来越多的证据表明,线粒体中的circRNA参与肿瘤细胞和TICs的自我更新。然而,迄今为止,线粒体编码的circRNA在肿瘤发生和TIC中的功能和调控机制尚不清楚。2023年1月,郑州大学生命科学学院朱平平教授课题组在Nature子刊Nature Communications发表文章mcPGK1-dependent mitochondrial import of PGK1 promotes metabolic reprogramming and self-renewal of liver TICs。作者鉴定了一种肝脏TICs中富集的线粒体编码的环状RNA——mcPGK1。其下调会损害肝脏TICs的自我更新,而过表达则相反。在调节机制上,mcPGK1通过抑制线粒体氧化磷酸化(OXPHOS)和促进糖酵解进而调节代谢,改变细胞内α-酮戊二酸和乳酸(Wnt/β-catenin激活和肝脏TICs自我更新的调节剂)的水平。此外,mcPGK1与TOM40的互作通过PGK1-PDK1-PDH轴从氧化磷酸化到糖酵解促进PGK1进入线粒体。文章解析了线粒体编码的circRNA调控线粒体功能、代谢和肝脏TICs自我更新的机制。mcPGK1在肝脏TICs中高表达首先,作者在肝脏TICs和非TICs中筛选出差异表达的6个mecircRNA,经过R酶消化、PCR和DNA测序鉴定均为circRNA。通过成球测试,作者选择mecirc8(mcPGK1)进行下一步分析。R酶消化验证mcPGK1稳定性,FISH验证其定位细胞质;mcPGK1在肝脏肿瘤中富集,并主要富集在线粒体外膜和基质中。mcPGK1的表达与临床分期、肿瘤体积、复发和生存相关。综上所述,线粒体编码的mcPGK1在肝脏TICs中稳定表达。图1.mcPGK1在肝脏TICs中高表达mcPGK1驱动肝脏TICs的自我更新作者评估了mcPGK1在肝脏TICs中的功能。敲低mcPGK1降低成球和增殖能力,过表达促进肝脏TICs的增殖和肿瘤生长。作者还针对线粒体mcPGK1构建了靶向纳米颗粒—shmcPGK1,在肝脏TICs上应用结果显示,成球能力受损。同样针对线粒体mcPGK1构建了靶向纳米颗粒—过表达mcPGK1,在肝脏TICs上应用结果与shmcPGK1相反。这些结果表明mcPGK1是肝脏TICs自我更新所必需的。图2. mcPGK1驱动肝脏TICs的自我更新mcPGK1改变OXPHOS到糖酵解的代谢过程线粒体的核心功能是能量代谢,作者评估了mcPGK1对OXPHOS和糖酵解的影响。在mcPGK1敲低的细胞中,OXPHOS活性增强,代谢产物增加,糖酵解活性减弱,而mcPGK1过表达则相反,这表明mcPGK1改变了OXPHOS转向糖酵解的代谢进程。作者随后测定了糖酵解和OXPHOS的两种主要代谢产物——乳酸和α-KG的表达。mcPGK1过表达情况下,乳酸迅速积累,但在mcPGK1敲低中没有;mcPGK1敲低的情况下,α-KG迅速积累,但在mcPGK1过表达中没有。这些数据表明mcPGK1改变了代谢过程。图3. mcPGK1改变OXPHOS到糖酵解的代谢过程Wnt信号调控mcPGK1的代谢变化来增强肝脏TICs功能乳酸促进肝脏TICs的自我更新和相关基因的表达,而α-KG具有相反的作用。二者可作为肝脏TICs的调控因子。作者又用FX-11(乳酸生成抑制剂,可抑制肝脏肿瘤的传播),验证了乳酸在肝脏TICs的作用。这些结果证实了代谢变化参与了肝脏TICs的生长。实验结果发现,乳酸和α-KG都以Wnt/β-catenin通路为靶点,而Wnt/β-catenin是影响肝脏TIC功能的关键信号通路。mcPGK1过表达增强的成球可被Wiki4或LF3抑制;mcPGK1过表达增加的乳酸也增强Wnt/β-catenin信号,而α-KG则相反,β-catenin(Wnt信号关键靶标)表达的结果也证明了这一结果。这些结果表明mcPGK1通过Wnt/β-catenin途径发挥作用。随后作者又用乳酸通过增强lnc-β-catm(可促进β-catenin的甲基化)表达促进β-catenin稳定,用α-KG通过H3K4me3去甲基化酶JARID1B抑制了β-catenin的转录,佐证了mcPGK1影响的乳酸和α-KG通过Wnt信号促进肝脏TICs生长。图4. Wnt/β-catenin介导乳酸和α-KG调控TICs生成mcPGK1与PGK1和TOM40互作,并促进PGK1与TOM40复合物互作circRNA Pulldwn被用于探索mcPGK1的分子机制。PGK1、TOM40和TOM70在下拉蛋白中,表明在肝脏TICs中与mcPGK1结合;WB也佐证这一结果。PGK1作为糖酵解和OXPHOS的关键调控因子,被选中用于机制探索,并再度验证出与mcPGK1互作。此外,作者发现,PGK1与TOM40/TOM70(可形成蛋白进入线粒体的通道)也存在互作,也就意味着mcPGK1同PGK1与TOM40/TOM70三者均有互作并发挥功能。不过,过表达mcPGK1可促进PGK1与TOM40/TOM70的结合,敲低mcPGK1则相反;作者突变了mcPGK1的不同位点验证了这种影响可能发生在特定位点(HR#2)。总而言之,mcPGK1与PGK1和TOM40互作,并促进PGK1与TOM40复合物互作。图5. mcPGK1促进PGK1与TOM40复合物的结合mcPGK1驱动PGK1进入线粒体PGK1在细胞质中表达,但在肿瘤发生过程中经常被转移到线粒体。作者发现,mcPGK1过表达促进PGK1进入线粒体,而敲低mcPGK1则相反;电镜的结果与之保持一致;并且敲除mcPGK1降低了PDH的磷酸化。这表示,mcPGK1-PGK1-PDK1介导的PDH磷酸化抑制了PDH将丙酮酸转化为乙酰辅酶a的功能,从而改变了从OXPHOS到糖酵解的代谢。最后作者评估了PGK1-PDK1-PDH轴在mcPGK1功能中的作用。纳米颗粒技术驱动PGK1进入线粒体后,可在很大程度上削弱mcPGK1敲低的作用;此外,过表达mcPGK1在抑制PDK1表达的条件下,成球的促进作用被限制,这表明mcPGK1驱动PGK1进入线粒体,然后发挥作用。综上表示,mcPGK1通过PGK1-PDK1-PDH途径驱动PGK1进入线粒体,抑制OXPHOS并促进糖酵解,从而促进肝脏TICs的生长。图6. mcPGK1促进PGK1进入线粒体原文链接:https://doi.org/10.1038/s41467-023-36651-5参考文献:[1] Guarnerio, J. et al. Oncogenic role of fusion-circRNAs derived from cancer-associated chromosomal translocations. Cell 165, 289–302 (2016).转载请联系邮箱授权:circRNA@163.com